Vertebrate brain theory
ISBN
978-3-00-064888-5
Monograph of Dr. rer. nat. Andreas Heinrich Malczan
4.5 The origin of the frontal cortex of vertebrates
With the development of signal divergence in the nucleus olivaris, the new signal class of extreme value-coded signals of the spinocerebellum was created. These were simultaneously digitized signals. They left the spinocerebellum and contacted the layers in the neural tube in which the commissure neurons of classes 4 and 5 were located. The class 5 neurons were located in the nucleus ruber and were responsible for the downward projection to the motoneurons, which has already been described.
But also (in the course of progressive evolution) new class 4 neurons could be
generated to which the axons of the cerebellum nuclei could dock. The class 4
neurons projected headwards. At the level of the thalamic level they contacted -
in analogy to the ancient rope ladder system - new class 4 projection neurons,
which in turn formed a new nucleus, the thalamic nucleus ventralis lateralis.
This was the beginning of the formation of a new, secondary turning loop. Their
input came from the spinocerebellum, and their output would again have to reach
the motor neurons of the neural tube or spinal cord. As is usual in the rope
ladder system, the axons of the upwardly projecting class 4 neurons of the new
thalamus had to dock with commissure class 3 neurons, which projected to the
motor part of the neural tube, where the motor spar was located in the rope
ladder system.
The class 3 neurons in turn contacted new class 5 projection neurons on the
motor side, which again projected downwards. The signal targets were the same:
the motoneurons of the muscles.
The new cortical turning structure above the thalamic level, which could not
have existed before the development of the spinocerebellum, because it processed
the output of the spinocerebellum, became the frontal cortex of vertebrates. Its
output was mainly motor and for the motor neurons of the muscles.
The main problem, however, was that due to the strong signal divergence there were significantly more signals than motor neurons. It was the same problem that had already been solved with the nucleus ruber or nucleus centromedianus. The solution was a strong signal convergence. A large amount of cortical input signals had to converge to a relatively small amount of output signals. In the nucleus ruber and in the nucleus centromedianus this led to the formation of magnocellular neurons, which formed their own nucleus part and managed the signal convergence.
For the new, secondary reversing loop, which received the output of the nucleus ventralis lateralis and ultimately directed it to the motor neurons, a convergence circuit was therefore urgently required if the output was to be used for motor control as before. The development of such a convergence circuit was on the evolutionary agenda.
In retrospect, we can state that in the vertebrate series, various approaches were taken to solve this convergence task, which ultimately led to the splitting of the vertebrates into reptiles, birds and mammals. In mammals, the secondary turning loop developed into the frontal cortex. The convergence circuit became the frontal lobe of the mammalian brain. This is located on the frontal outer side of the brain.
In reptiles and birds, the convergence circuit remained inside the ventricular cavity and formed the dorsal ventricular crest (DVR). This convergence circuit increasingly filled the ventricular space until further spatial growth was no longer possible. Especially the strong head skeleton of the reptiles prevented further spatial expansion. Thus the reptiles simply stopped at a relatively low level in their brain development.
The birds were able to shift some of the necessary convergence circuitry to the outer surface of the brain, but retained the organizing principle of the DVR. The new convergence structure in the birds was called the hyperpallium.
Therefore, we have to consider the further course of brain evolution for reptiles, birds and mammals separately from now on, although, for example, with regard to the basal ganglia and the cerebellum, there are great analogies with regard to neuronal circuits.
Theorem of the origin of the frontal cortex of mammals
With the development of the spinocerebellum and the onset of signal divergence in the nucleus olivaris,the thalamic nucleus ventralis lateralis became the target structure for afferent projection. Like all cortical turning loops, the frontal cortex breaks down into a sensory input part and a motor output part. The input part receives in the class 4 neurons the signals coming from the cerebellum via the thalamus and switches them to class 3 commissure n
Additionally, there are class 6 mean neuronsthat project to mean structures and class 1 neuronsthat receive mean excitations and activate neighboring neurons of different classes. This structure of the frontal cortex was developed only in mammals, in reptiles and birds there were parallel developments with other neural circuits.
The topology in the frontal cortex
The Frontalcortex can be compared to a concave mirror, which receives the input on the sensor side and "reflects" it to the motor side, where it is passed down as output. This also results in signal reflection.
A special feature of the motor side is the phenomenon of signal convergence, which cancels out the original signal divergence in the nucleus olivaris before the converged signals are fed to the motor system.
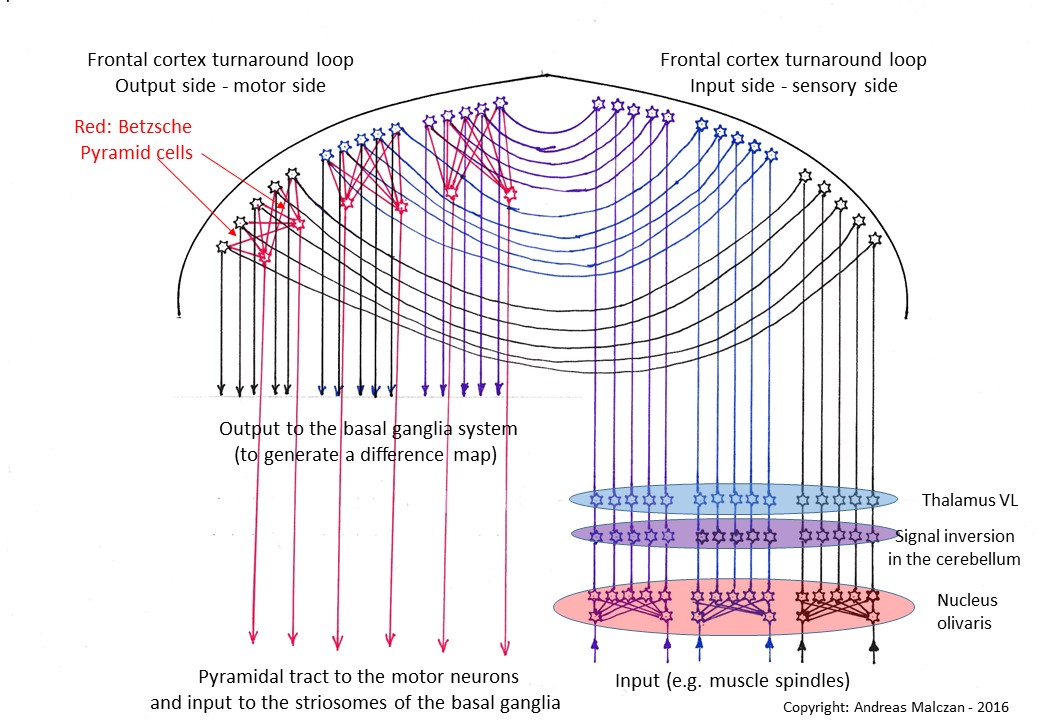
Figure
41 - The Frontalcortex as a New Turning Structure
and Convergence System
Theorem of Betz's giant pyramid cells
The signal divergence caused by the nucleus olivaris was reversed in the frontal cortex by Betz's giant pyramid cells. Their projection axons form the pyramidal tractthat projects to the motor neurons of the spinal cord.
The Betz cells solved the first problem that the motor frontal cortex had to solve. There were significantly fewer motor neurons in the spinal cord than output signals in the frontal cortex. The muscle spindle signals had been subjected to a strong signal divergence in the nucleus olivaris, which now had to be reversed in the frontal cortex as well as in the nucleus ruber or in the nucleus centromedianus. This is how the type of neuron was created in the frontal cortex, which uses a huge dendrite tree to subject the incoming signals to a strong signal convergence. These integration neurons are today called Betzsche cells. They are giant pyramid cells in the fifth layer of the cortex and they project to the conscious motor neurons. And they are located on the output side of the frontal cortex and belong to neuron class 5.
The axons of these Betz's giant pyramid cells projected directly to the motor
neurons. To do this, they had to move downwards and, among other things, find a
path through the structure known as striatum. Thus, the striatum was divided
into the caudate nucleus and the putamen. The projection of Betz' cells in
mammals is also called pyramidal trajectory.
In the above illustration, the layer structure has been neglected, as only the
principle circuit diagram has been shown.
There is a great analogy between the cortex layer of the frontal cortex and the layers of the different neuron nuclei of the vertebrate brain system. We have already observed such a neuron layer in the nucleus olivaris, which is a functional structure open on one side. Likewise, the cerebellar nuclei became such two-dimensional structures. The cerebellum layer also formed a surface. Now, above the thalamic nucleus ventralis lateralis, such a flat structure also developed.
And like all surfaces, this layer again formed a body model of the associated receptors or motor neurons, ordered by modalities and body segments.
It contained - just like the rope ladder system before - both the receiving neurons and the descending neurons. This area broke into two parts, just as the rope ladder system had previously been divided into two parts. One half was responsible for the input and contained the receiving neurons. The other half was responsible for the output and contained the descent neurons, i.e. the output neurons.
The input and output neurons formed pairs of neurons, with each input neuron contacting the corresponding output neuron via a class 3 mediating neuron. This neuron area formed the frontal lobe of the primordial brain.
The original goal of the signals ascending from the spinal cord, which consisted in motor control, was retained for the frontal lobe. Its output signals, like the original cerebellum output, were intended to control the motor neurons.
Due to the unfolding of the neural tube as a result of the necessity to supply the externally running axons of the nucleus olivaris to the developing cerebellum, the spatial arrangement of grey and white matter was reversed in the frontal lobe. The neurons of the developing frontal cortex sought the proximity of the outer surface of the brain, which bordered on the outer cerebrospinal fluid space. Thus, the cell nuclei were located in the outer surface of the frontal cortex, while only the interior remained for the axons to use.
An axon guide was used in the frontal lobe, which may be described here as an inversion mirroring and which led to the topology inversion. The outer surface of the frontal lobe, which did not show any convolution at that time, behaved to the signals in a similar way to the surface of a concave mirror. On the input side, the ascending axons - running roughly parallel to each other - met their receiving neurons. Their axons reversed direction and ran downwards, following circular paths towards the output side. Here, too, the axons ran next to each other in a well-ordered manner without crossing each other. This is how they reached the output neurons on the output side of the frontal cortex.
The advantage of this axon course was the possibility that not only the signals of the muscle spindles could contact the motor output neurons, but also those of the other signals. In the meantime, there were enough trunk and head senses that already achieved motor effects by reacting with the muscle spindle signals in the torus semicircularis or the tectum opticum, for example. And because the associated signals did not end in these association areas, but continued on their own axons in the cortical turning loop and reached the cerebellum via the nucleus olivaris, there was also non-motor cerebellum output. But because of the synaptic coupling to the motor signals in the association areas of the torus and the tectum, these signals were now signal-related to the motor signals.
The resulting signal relationship enabled the sensory signals to also affect the motor output neurons in the frontal cortex by projecting from the sensory input surface of the frontal cortex via axons into the motor output surface of the frontal cortex. This was because one task of the trunk and head senses was the motor control of the living being. So there were also projections from the sensory input surfaces of the Frontalcortex into the motor output surfaces. In this way the frontal cortex could take over the tasks that had previously been carried out in the torus semicircularis or the tectum, thus gaining control over these subsystems and becoming their superior system. Subsequently, a reduction of the tasks of the torus and the tectum occurred.
Here it also becomes clear that only the signals of the muscle spindles formed the actual output signals of the frontal cortex. Because all input areas of the sensory frontal cortex ultimately projected onto this motor frontal cortex.
Like all bilateral subsystems, the frontal cortex is duplicated and thus exists on each of the two sides of the body.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan