Gehirntheorie der Wirbeltiere
ISBN
978-3-00-064888-5
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
4.5 Die Entstehung des Frontalcortex der Wirbeltiere
Mit der Herausbildung der Signaldivergenz im Nucleus olivaris entstand die neue Signalklasse der extremwertcodierten Signale des Spinocerebellums. Diese waren gleichzeitig digitalisierte Signale . Sie verließen das Spinocerebellum und kontaktierten im Neuralrohr die Schichten, in denen sich die Kommissurneuronen der Klassen 4 und 5 befanden. Die Neuronen der Klasse 5 befanden sich im Nucleus ruber und waren für die Abwärtsprojektion zu den Motoneuronen zuständig, die bereits beschrieben wurde.
Aber ebenso konnten (im Verlaufe der fortschreitenden Evolution) auch neue Neuronen der Klasse 4 generiert werden, an denen die Axone der Cerebellumkerne andocken konnten. Die Neuronen der Klasse 4 projizierten kopfwärts. Auf der Höhe der thalamischen Etage kontaktierten sie - in Analogie zum uralten Strickleitersystem - neue Projektionsneuronen der Klasse 4, die ihrerseits einen neuen Kern, den thalamischen Nucleus ventralis lateralis bildeten. Dies war der Beginn der Entstehung einer neuen, sekundären Wendeschleife. Ihr Input entstammte dem Spinocerebellum, und ihr Output müsste wieder die Motoneuronen des Neuralrohrs bzw. des Rückenmarks erreichen. Wie beim Strickleitersystem üblich, mussten die Axone der aufwärts projizierenden Neuronen der Klasse 4 des neuen Thalamus an Kommissurneuronen der Klasse 3 andocken, die zum motorischen Teil des Neuralrohrs projizierten, wo sich beim Strickleitersystem der motorische Holm befand.
Die Neuronen der Klasse 3 ihrerseits kontaktierten auf der motorischen Seite neue Projektionsneuronen der Klasse 5, die wieder abwärts projizierten. Die Signalziele waren die gleichen: die Motoneuronen der Muskeln.
Die neue cortikale Wendestruktur oberhalb der thalamischen Etage , die es vor der Entwicklung des Spinocerebellums gar nicht gegeben haben kann, weil sie den Output des Spinocerebellums verarbeitete, wurde der Frontalcortex der Wirbeltiere . Sein Output war vorwiegend motorisch und für die Motoneuronen der Muskeln bestimmt.
Das Hauptproblem bestand aber darin, dass es durch die starke Signaldivergenz deutlich mehr Signale als Motoneuronen gab. Es war das gleiche Problem, welches schon beim Nucleus ruber oder Nucleus centromedianus gelöst worden war. Die Lösung bestand in einer starken Signalkonvergenz . Eine große Menge von cortikalen Inputsignalen musste auf eine relativ kleine Menge von Outputsignalen konvergieren. Im Nucleus ruber und im Nucleus centromedianus führte dies zur Herausbildung magnocellularer Neuronen , die einen eigenen Kernanteil bildeten und die Signalkonvergenz bewerkstelligten.
Für die neue, sekundäre Wendeschleife, die den Output des Nucleus ventralis lateralis entgegennahm und letztlich zu den Motoneuronen leitete, war also eine Konvergenzschaltung dringend erforderlich, wenn der Output wie bisher zur motorischen Steuerung eingesetzt werden sollte. Die Entwicklung einer solchen Konvergenzschaltung stand auf der Tagesordnung der Evolution.
Im Nachhinein können wir feststellen, dass in der Wirbeltierreihe zur Lösung dieser Konvergenzaufgabe verschiedene Lösungswege beschritten wurden, die letztlich zur Aufspaltung der Wirbeltiere in die Reptilien, die Vögel und die Säugetiere führten. Bei den Säugetieren entwickelte sich die sekundäre Wendeschleife zum Frontalcortex . Die Konvergenzschaltung wurde zum Frontallappen des Säugergehirns. Dieser befindet sich an der vorderen Außenseite des Gehirns.
Bei Reptilien und Vögeln blieb die Konvergenzschaltung im Inneren des Ventrikelraumes und bildete den dorsalen ventrikulären Kamm (DVR ). Diese Konvergenzschaltung füllte den Ventrikelraum zunehmend aus, bis ein weiteres räumliches Wachstum nicht mehr möglich war. Speziell das starke Kopfskelett der Reptilien verhinderte eine weitere räumliche Ausdehnung. So blieben die Reptilien in ihrer Gehirnentwicklung auf einer relativ niedrigen Stufe einfach stehen.
Die Vögel konnten einen Teil der nötigen Konvergenzschaltung auf die Außenfläche des Gehirns verlagern, behielten jedoch das Organisationsprinzip des DVR bei. Die neue Konvergenzstruktur wurde bei den Vögeln das Hyperpallium .
Daher müssen wir den weiteren Werdegang der Gehirnevolution für Reptilien, Vögel und Säuger von nun an getrennt betrachten, obwohl etwa hinsichtlich der Basalganglien und des Cerebellums große Analogien hinsichtlich der neuronalen Schaltungen auftreten.
Theorem der Entstehung des Frontalcortex der Säugetiere
Mit der Entstehung des Spinocerebellums und der einsetzenden Signaldivergenz im Nucleus olivaris entstand als Zielstruktur für die afferente Projektion der thalamische Nucleus ventralis lateralis , der die Cerebellumsignale entgegennahm und einer neuen cortikalen Wendeschleife zustellte, die als Frontalcortex bezeichnet wird. Dieser zerfällt - wie alle cortikalen Wendeschleifen - in einen sensorischen Inputteil und einen motorischen Outputteil. Der Inputteil empfängt in den Neuronen der Klasse 4 die vom Cerebellum über den Thalamus kommenden Signale und schaltet sie auf Kommissurneuronen der Klasse 3 um, die zum motorischen Outputteil ziehen und dort Konnektivneuronen der Klasse 5 kontaktieren, die abwärts projizieren.
Zusätzlich gibt Mittelwertneuronen der Klasse 6 , die zu Mittelwertstrukturen projizieren sowie Neuronen der Klasse 1 , die Mittelwerterregungen empfangen und benachbarte Neuronen verschiedener Klasse aktivieren.
Diese Struktur des Frontalcortex wurde nur bei Säugetieren entwickelt, bei Reptilien und Vögeln gab es parallele Entwicklungen mit anderen neuronalen Schaltungen.
Die Topologie im Frontalcortex
Der Frontalcortex ist hinsichtlich des Signalverlaufs vergleichbar mit einem Hohlspiegel , der auf der Sensorikseite den Input empfängt und ihn zur motorischen Seite -reflektiert-, wo er als Output nach unten weitergegeben wird. Dadurch tritt auch eine Signalspiegelung auf.
Als Besonderheit der motorischen Seite tritt das Phänomen der Signalkonvergenz auf, welche die ursprüngliche Signaldivergenz im Nucleus olivaris wieder aufhebt, bevor die konvergierten Signale der Motorik zugeführt werden.
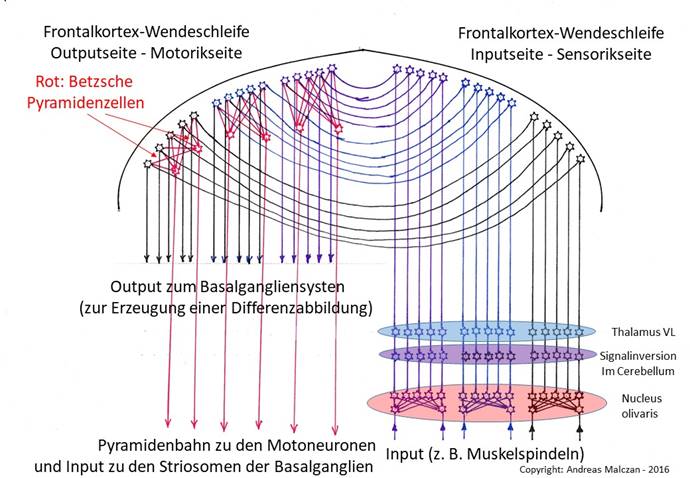
Abbildung 41 - Der Frontalcortex als neue Wendestruktur und Konvergenzsystem
Theorem der Betzschen Riesenpyramidenzellen
Die durch den Nucleus olivaris verursachte Signaldivergenz wurde im Frontalcortex durch die Betzschen Riesenpyramidenzellen wieder aufgehoben. Ihre Projektionsaxone bilden die Pyramidenbahn , die zu den Motoneuronen des Rückenmarks projiziert.
Die Betzschen Zellen lösten das erste Problem, welches der motorische Frontalcortex zu lösen hatte. Es gab deutlich weniger Motoneuronen im Rückenmark als Outputsignale im Frontalcortex. Die Muskelspindelsignale waren im Nucleus olivaris einer starken Signaldivergenz unterworfen worden, die nun im Frontalcortex ebenso rückgängig gemacht werden musste wie im Nucleus ruber oder im Nucleus centromedianus. So entstand also auch im Frontalcortex jene Neuronenart, die mit einem riesigen Dendritenbaum die eintreffenden Signale einer starken Signalkonvergenz unterzieht. Diese Integrationsneuronen werden heute als Betzsche Zellen bezeichnet. Es sind Riesenpyramidenzellen in der fünften Schicht des Cortex und sie projizieren zu den bewussten Motoneuronen. Und sie befinden sich auf der Outputseite des Frontalcortex und gehören der Neuronenklasse 5 an.
Die Axone dieser Betzschen Riesenpyramidenzellen projizierten direkt zu den Motoneuronen. Hierbei mussten sie abwärts ziehen und sich unter anderem einen Weg durch diejenige Struktur bahnen, die als Striatum bezeichnet wird. So wurde das Striatum zweigeteilt in den Nucleus caudatus und das Putamen . Die Projektion der Betzschen Zellen wird bei den Säugern auch als Pyramidenbahn bezeichnet.
In der obigen Darstellung ist der Schichtenaufbau vernachlässigt worden, da nur das Prinzipschaltbild dargestellt wurde.
Es gibt eine große Analogie zwischen der Cortexschicht des Frontalcortex und den Schichten der verschiedenen Neuronenkerne des Gehirnsystems der Wirbeltiere. Eine solche Neuronenschicht beobachteten wir ja bereits beim Nucleus olivaris, der ein sachartiges, an einer Seite offenes Gebilde darstellt. Ebenso wurden die Kleinhirnkerne zu solchen flächigen Strukturen. Auch die Cerebellumschicht bildete eine Fläche. Nun entstand oberhalb des thalamischen Nucleus ventralis lateralis ebenfalls eine solche flächige Struktur.
Und wie alle Flächen bildete diese Schicht wieder ein nach Modalitäten und Körpersegmenten geordnetes Körpermodell der zugehörigen Rezeptoren bzw. Motoneuronen.
Sie enthielt - ebenso wie zuvor das Strickleitersystem - sowohl die Empfangsneuronen als auch die Abstiegsneuronen. Hierbei zerfiel diese Fläche in zwei Teile, so wie zuvor auch das Strickleitersystem zweigeteilt war. Eine Hälfte war für den Input zuständig und enthielt die Empfangsneuronen. Die andere Hälfte war für den Output zuständig und enthielt die Abstiegsneuronen, also die Outputneuronen.
Die Input- und Outputneuronen bildeten Neuronenpaare, wobei jedes Inputneuron über ein vermittelndes Neuron der Klasse 3 das zugehörige Outputneuron kontaktierte. Diese Neuronenfläche bildete den Frontallappen des Urhirns.
Das ursprüngliche Ziel der vom Rückenmark aufsteigenden Signale, welches in der motorischen Steuerung bestand, blieb für den Frontallappen erhalten. Seine Outputsignale sollten, ebenso wie der ursprüngliche Cerebellumoutput, die Motoneuronen ansteuern.
Durch die Auffaltung des Neuralrohrs infolge der Notwendigkeit, die außen verlaufenden Axone des Nucleus olivaris dem entstehenden Cerebellum zuzuführen, wurde beim Frontallappen die räumliche Anordnung von grauer und weißer Substanz umgekehrt. Die Neuronen des entstehenden Frontalcortex suchten die Nähe der Gehirnaußenfläche, die an den äußeren Liquorraum grenzte. So befanden sich die Zellkerne in der Außenfläche des Frontalcortex, während den Axonen nur das Innere zur Benutzung verblieb.
Im Frontallappen wurde eine Axonführung verwendet, die hier als Inversionsspiegelung bezeichnet werden möge und die zur Topologieumkehr führte. Die Außenfläche des Frontallappens, die damals noch keine Faltung aufwies, verhielt sich zu den Signalen ähnlich wie die Fläche eines Hohlspiegels . An der Inputseite trafen die aufsteigenden Axone - etwa parallel zueinander verlaufend - auf ihre Empfangsneuronen. Deren Axone kehrten die Richtung um und verliefen nach unten, wobei sie auf kreisähnlichen Bahnen in Richtung der Outputseite verliefen. Auch hierbei verliefen die Axone wohlgeordnet nebeneinander, ohne sich zu kreuzen. So erreichten sie die Outputneuronen der Outputseite des Frontalcortex.
Der Vorteil bei diesem Axonverlauf bestand in der Möglichkeit, dass nicht nur die Signale der Muskelspindeln die motorischen Outputneuronen kontaktieren konnten, sondern auch die der anderen Signale. Denn in der Zwischenzeit gab es genügend Rumpf- und Kopfsinne, die bereits motorische Wirkungen erzielten, indem sie etwa im Torus semicircularis oder im Tectum opticum mit den Muskelspindelsignalen reagierten. Und weil die zugehörigen Signale nicht in diesen Assoziationsgebieten endeten, sondern auf eigenen Axonen in der cortikalen Wendeschleife weiterzogen und über den Nucleus olivaris auch das Cerebellum erreichten, gab es auch nichtmotorischen Cerebellumoutput. Doch wegen der synaptischen Kopplung zu den motorischen Signalen in den Assoziationsgebieten des Torus und des Tectums waren diese Signale nun signalverwandt mit den motorischen Signalen.
Die so entstandene Signalverwandschaft ermöglichte es den sensorischen Signalen, im Frontalcortex ebenso auf die motorischen Outputneuronen einzuwirken, indem sie von der sensorischen Inputflächen des Frontalcortex über Axone in die motorische Outputfläche des Frontalcortex projizierten. Denn eine Aufgabe der Rumpf- und Kopfsinne war die motorische Steuerung des Lebewesens. So gab es also auch Projektionen von den sensorischen Inputflächen des Frontalcortex in die motorischen Outputflächen . Auf diese Art konnte der Frontalcortex die Aufgaben übernehmen, die zuvor im Torus semicircularis oder im Tectum realisiert worden waren, gewann so die Oberhoheit über diese Subsysteme und wurde deren übergeordnetes System. In der Folge trat eine Reduktion der Aufgaben des Torus und des Tectums auf.
Hier wird auch klar, dass nur die Signale der Muskelspindeln die eigentlichen Outputsignale des Frontalcortex bildeten. Denn alle Inputgebiete des sensorischen Frontalcortex projizierten letztlich auf diesen motorischen Frontalcortex.
Wie alle bilateralen Subsysteme ist auch der Frontalcortex doppelt vorhanden und existiert somit auf jeder der zwei Körperseiten.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan