Vertebrate brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-064888-5
4.4The splitting of the neural tube during cerebellum development
We first consider the spatial arrangement of neurons and their axons in the neural tube. On the inside, the cell bodies of the neurons were arranged to form the grey matter, while the axons ran on the outside. In the middle was the ventricular space, one of its tasks was to secure the metabolic processes of the neurons, the supply of nutrients and the removal of metabolic products.
Since in the system described so far (almost) all signals of the nucleus ruberreached the cerebellumvia the nucleus olivaris , this number of signals was also quite large in the early phase. Just imagine that for each descending axon of the upper neural tube section containing the output of the primordial brain, there was also an axon that reached the cerebellum via the nucleus ruber and the nucleus olivaris.
With the onset of signal divergence in the nucleus olivaris, this axon number was greatly increased in the direction of the cerebellum. As the degree of divergence in the nucleus olivaris increased, the number of climbing fibres moving towards the cerebellum increased.
These very numerous axons had to travel from the outside of the neural tube - they came there from the olive pit on the opposite side - to the inside, because that is where the Purkinje cells were located. As a result, the neural tube on the cerebellum side bent apart like a slit to allow passage for the many axons that were moving towards the cerebellum. As the number of axons passing through widened, the gap widened both in width and height.
The resulting opening had the appearance of a rhombus placed on top. Through this opening, the axons located on the outside of the neural tube moved inwards to the cerebellum neurons located on the inside. At the same time, an opening was created in the spinal canal. The spinal fluid was able to flow out through this rhomboid gap and form a ventricle at the cerebellum, which can still be found in all vertebrates today.
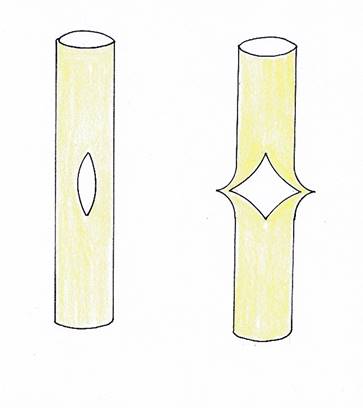
Abbildung40- Aufspaltung des Neuralrohrs
In the second expansion phase of the cerebellum we observe an explosive increase in the number of Purkinje cells. This led to an enormous expansion of the cerebellum, which now became approximately spherical, while still consisting of two halves. Although the cell bodies of the nerve cells in the neural tube were generally arranged inside, while the associated axons were arranged outside, this rule was reversed. In the neural tube itself, there was no longer sufficient space for the cerebellum, so that the enlarging structure of the cerebellum was pushed outwards through the resulting rhomboid-shaped gap, creating a kind of spatial protrusion of the neural tube. Thus the cerebellum had migrated from the inside of the neural tube to the outside. The section of the neural tube that is at the level of the developing cerebellum is called the rhombic brain (rhombencephalon).
The described unfolding of the neural tube had consequences for the position of the posterior and anterior horn at this height. If the hitherto hind horn was at the rear, i.e. backwards, while the front horn was at the front, i.e. nose-wards, the unfolding led to a displacement of both, so that they were located approximately side-by-side. Since the opening of the neural tube took place on the rear side, i.e. between the left and right posterior horn, the left posterior horn moved to the left and the right posterior horn to the right. Likewise, the cortical turning loop was also migrated, in which the posterior horn axons moved upwards and made the change of direction from the sensory to the motor side in order to move to the motor neurons in the anterior horn. The posterior horn was now no longer at the rear but on the outside of the anterior horn (shoulder direction). Thus, the axons of the posterior horn ran from the left and right outside to the inside. The cortical reversible structure had turned by about 90 degrees.
This twisted turning loop later became the temporal lobe of the brain. It lies laterally on the left or right side of the brain and is present twice because of the bilateral symmetry. The temporal lobe was later pushed aside by the developing frontal lobe.
Between the two temporal lobes a small part of the turning loop was left, which could no longer follow this unfolding because it was already ancient and too conservative from an evolutionary point of view. It was responsible for the signals of the elementary life-support system. In modern vertebrates, this remaining part forms the island cortex of the brain .
There is sufficient evidence for this thesis of the author. Even today the processing of all hair cell senses takes place in the temporal lobe.This is the case, for example, with the sense of hearing, whose function is made possible by the hair cells.
We also find the olfactory and limbic systemin the temporal lobe, as well as a part of the original spinal canal, which becamethe ventricle system in the cortical turning loop.
Another consequence of the unfolding of the neural tube was the opening of the previously closed ventricular space of the neural tube. The opening allowed the ventricular fluid to escape and to flow around the previously formed structure of nerve cells, axons, dendrites and other components from the outside. It can be assumed that step by step a kind of bubble was formed, which surrounded the previous system from the outside. Thus it happened that the entire central nervous system was surrounded by fluid both from the inside and the outside.
This had another consequence. The neurons of the neural tube always sought proximity to the ventricular wall in terms of their spatial position. Now there was an additional, outer ventricular wall, which initially surrounded only the developing cerebellum, later also the whole developing brain. And the cell bodies of the neurons still sought their spatial proximity. This led to a strange result. While the grey matterin the neural tube and in the later spinal cord is located inside, the grey matter of the cerebellum and the cerebrum is generally located outside. This is because the associated neurons still seek contact or proximity to the ventricular wall, but this wall has now been turned outwards by the unfolding of the neural tube.
And while in the neural tube newly formed neurons were always formed on the inside of the neural tube during evolution, in the cerebellum and in the cerebrum it was exactly the opposite. There, the evolutionarily older neurons are located on the inside and the younger ones on the outside.
Through the opening formed by the neural tube during the unfolding process, the ventricular fluid came out as described above and caused the entire central nervous system to be stored in a fluid bubble. This was extremely advantageous for such a sensitive organ. The ventricular fluid acted like a shock absorber and protected the developing brain from strong impacts. Thus it fulfilled not only chemical and biological but also physical tasks.
In the course of evolution, however, the axon bundles in the neural tube also split up to transport signals. This probably began in the upper section, because the growth of the head simply made more space available there. In principle, the axons began to arrange themselves according to modalities. This process initially took place in the uppermost levels of the neural tube. Later on, this division according to modalities was partially transferred to the other levels of the brain system. First, however, axon structures, which we will refer to as modality rings, developed in the uppermost levels of the neural tube.
Theorem of the splitting of the modality rings in the head region of the neural tube
Each modality ring of the neural tube in the trunk area consists of two sensory and two motor quarter rings. One sensory and one motor modality quarter-ring each represent one half of the body.
Between the third and second segment, the nested sensory modality quarter rings are split into adjacent sensory modality bands. A splitting of the motor modality quarter-rings was not performed. The projection neurons of class 4 of each modality ring of the third segment project to the projection neurons of class 3 and 4 of the second segment, whereby their axons abandoned the previous arrangement in modality rings and in the second segment the modalities are arranged next to each other in a strip-like manner. Each modality now forms a kind of rectangle in cross-section. The previous organisation of neurons in a sensory centre is abandoned. The original sensory center decays into modality centers .
In fact, there are now as many sensory individual nuclei as there are modalities. The individual nuclei are arranged in a row next to each other and form a structure that is called thalamus in vertebrates.
Since the splitting affects both the left-sided and the right-sided sensory modality quarter ring, the living being has two thalami.
The individual thalamus nuclei project into the first segment, as is also usual in the rope ladder nervous system. However, the splitting of the sensory center of the second segment into a number of adjacent modality centers has consequences for the arrangement of the neurons in the first segment.
First of all, it should be noted that the space requirement is increasing enormously. The adjacent modality centers - which form the later thalamus - require more space. The neural tube therefore inflates at the head area. The space is needed on both the left and the right, because each half of the body has such a structure. Thus, in the neural tube, a strong bulge is created on both the left and right side, while the olfactory middle part is preserved and bulges forward. One could speak of a three-part structure, a three-bubble stage.
Simplified we think of the modality stripes per modality of constant width. Placed side by side, a kind of rectangular surface is created, but it is curved. The easiest way to describe the curved surface is as the surface of a quarter cylinder. It consists of the individual modality strips. The width of the stripes depends on the number of receptors belonging to the modality in question.
Inside a modality strip, the axons of the class 4 projection neurons run from the second segment and meet class 4 neurons on the outer surface of the strip, which originally belonged to the sensory core of the first segment. Now, after splitting the sensory core into separate modality nuclei, these neurons become the input neurons of the first segment. Since there is no further segment above them, they mutate into interneurons. These class 4 receiving neurons of the first segment lie in a strip-like curved surface that belongs to a quarter cylinder. This inevitably results in a well-ordered arrangement of these neurons. The lowest segment, i.e. the tail segment, is represented as the uppermost horizontal stripe on the surface of the quarter cylinder. The receptor signals of the receptors of a particular modality arrive first, so to speak, in the cylindrical surface and contact the class 4 neurons there, which are the first to be encountered. The receptors of the second segment from below send their signals to the second horizontal strip below. The third segment from below projects into the third stripe, and so on. The uppermost segment, in which this modality is used, projects into the lowest strip of the cylindrical surface. This is because the class 4 projection axions of the lower segments are more inside the neural tube, whereas the upper ones are outside. Thus, they arrive at the upper cortical level and are also arranged in this way on its cortical outer surface.
Since we call the total area, in which the afferent projection neurons of the second segment end and transfer their excitation to the input neurons of class 4 there, a cortex, we can summarize the presented algorithms regarding the topological mapping.
Theorem of cortical well-being by segments
The cortex of the earliest vertebrates is not only well-ordered by modalities, but each modal stripe is well-ordered by body segments. The tail segment is the topmost segment strip of each modality, the head segment the lowest. Thus the cortex becomes a body model of the living being for each modality.
With the formation of new body components such as jaws, fins, limbs, etc., new building blocks are added to the existing topological body model of each modality. It must be noted here that this also led to the formation of new input and output cores. Thus, besides the nucleus gracilis and cuneatus and the nucleus ruber, further nuclei of this type were created. Already here the body image decays into a separate part representing the upper half and one that is assigned to the lower half. A middle part for the head receptors is added.
The new input cores are not discussed further in this monograph, but they do exist in reality and will have adopted many signal processing algorithms.
Outlook:
The splitting of the modalities first began in the thalamic plane. Over the course of millions of years, deeper segments were also included. In mammals, the splitting can already be detected in the spinal cord, where axons of the same modality form separate fiber bundles. This was made possible because the cortex took over the tasks previously performed by the connective tissue in the original cord ladder system of each half of the body: to form the autonomic apparatus.
One consequence of the splitting of the modalities was the formation of the different brain lobes, each of which is responsible for one modality group.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan