Vertebrate brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-064888-5
4.3The second expansion phase of the original Pontocerebellum
With the strong increase in the number of receptors in the course of evolution, it became more advantageous to summarize the cortical mean excitations before they were evaluated. Therefore, several class 6 cortical mean neurons converged to a common mean neuron of the subthalamic nucleus. This represented the mean nucleus of the thalamic floor.
Originally, each cortex cluster projected via its mean neuron into the subthalamic nucleus. Now, however, a group of adjacent cortex clusters projected into such a neuron.
If one thinks of the initially worm-shaped creature arranged vertically, so that the head is on top and the tail is below, the cortex as a surface had a similar, but much smaller shape. It was also divided into modality strips and segment strips. The nucleus subthalamicus as a body model of the cortical modalities had an analogous arrangement. For averaging, it was advantageous to keep the modality boundaries and segment boundaries. Therefore, only clusters were combined to a common mean value, which formed a cluster group horizontally next to each other to form a modality.
Definition: degree of convergence of the subthalamic nucleus
If the mean neurons of a cluster group of n adjacent cortex clusters project into a common output neuron of the subthalamic nucleus, then we refer to the number n as the degree of convergence.
The degree of convergence is thus the number of cortex clusters that project into a common output neuron of the subthalamic nucleus.
After the convergence of the cortical mean signals to the neurons of the nucleus subthalamicus, the cerebellum cortex of the pontocerebellum was still a cortex model with respect to the granule cells. However, the number of output neurons of the subthalamic nucleus that projected into the nucleus olivaris had significantly decreased - in relation to the number of corteclusters.
Theorem of cluster groups in the nucleus subthalamicus
Each cluster group X, consisting of adjacent cortex clusters, is represented in the subthalamic nucleus by a common mean neuron.
Theorem of the nucleus subthalamicus as a mean value body model
The nucleus subthalamicus represents a coarsened body modelof the cortex cortex in which each cortical cluster group is represented by a mean neuron.
The convergence of the mean value signals in the nucleus subthalamicus was certainly useful. Since each mean nucleus exchanged data with the mean nuclei of other segments, a reduction of the signal-carrying axons was material-saving. If one neuron failed, averaging was only slightly disturbed, since the other axons also provided mean values. On the other hand, the degree of convergence must not be too great, because otherwise the assignment of an average value to the signal-generating part of the body would no longer be precise enough.
We postulate here that the mean value model of the subthalamic nucleus reflected the segment structure of the body. Therefore, of the incoming axons, only those that represented signals from the same body segment were combined in a common output neuron. Thus, the dendrites of the output neurons ran approximately parallel to each other to the left and right when the head was on top and the tail was at the bottom in the body model. This projection also took into account the modalities that were kept strictly apart.
This signal convergence was reversed in the nucleus olivaris by an opposite signal divergence. This is because parallel to the signal convergence in the subthalamic nucleus, a signal divergence occurred in the olivaric nucleus. Here, the failure of a single neuron in the nucleus olivaris caused a sensitive disturbance of motor function. Therefore, an increasingly stronger signal divergence took place in the nucleus olivaris. Two input signals were therefore distributed to n output signals. This principle was also applied to the mean value regions of the nucleus olivaris, which projected into the Pontocerebellum.
If two adjacent cluster groups C1 and C2 (of the same segment) projected into the subthalamic nucleus and the striosome system generated two climbing fibre signals fA and fB for the Pontocerebellum, these signals were received by two adjacent input neurons of the olivar nucleus.
We assume that these input signals were divergently distributed in the nucleus olivaris to a group of output neurons arranged in a row. As evolution progressed, their number increased.
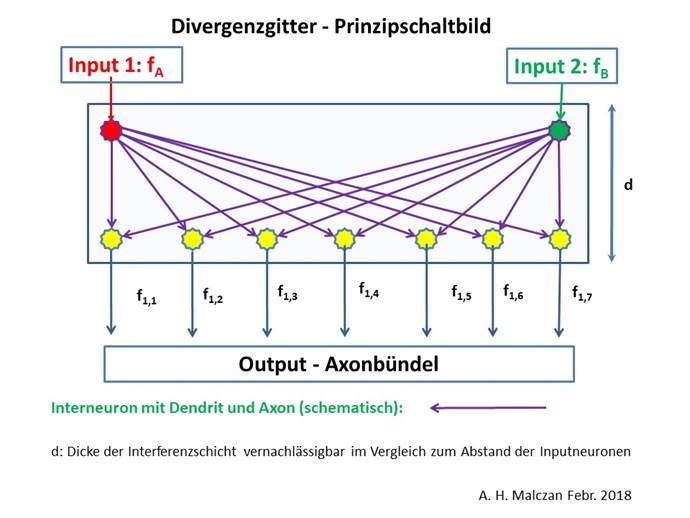
Abbildung 37 - Signaldivergenz im Nucleus olivaris
We repeat here an earlier definition analogously:
Definition: Degree of divergence in the nucleus olivaris
If two input signals fA and fB are distributed by signal divergence in the nucleus olivaris to a row of n output neurons, we refer to the number n as the degree of divergence of the nucleus olivaris.
We now assume that in the course of evolution the degree of convergence in the subthalamic nucleus and the degree of divergence in the olivaris nucleus were approximately equal.
Theorem of the inversion of signal convergence in the nucleus dentatus
In the nucleus dentatus the signal convergence of the nucleus olivaris was reversed by an approximately equal signal divergence.
The distribution of the climbing fibre signals is achieved by parallel divergence. The derived climbing fibers arrived in the Pontocerebellum and were again distributed by sequential divergence to several Purkinj groups arranged one behind the other.
Thus, in the Pontocerebellum, n climbing fiber signals were again available for a cluster group of n cortex clusters. Since both input partners fA and fB were higher frequency, the superposition result in the divergence lattice of the nucleus olivaris was also higher frequency despite the exponential attenuation. This was important for the use of these tetanic signals for long-term depression and long-term potentiation in the Pontocerebellum. But this also required parallel fiber signals.
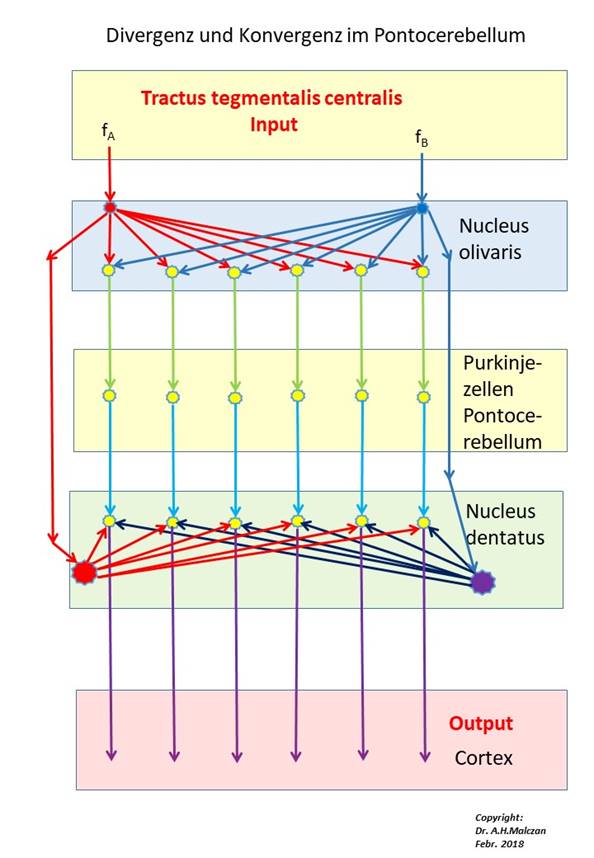
Abbildung 38 - Signaldivergenz und Signalkonvergenz im Pontocerebellum
In the figure above only the climbing fiber input to the Purkinje cells is shown, the signal inflow via the parallel fibers is (temporarily) omitted. The fire rates fA and fB do not only reach the two input neurons of the nucleus olivaris, but also the assigned neurons of the nucleus dentatus via the tractus tegmentalis centralis - after transformation into a tetanic climbing fiber signal with pauses in oscillation.
Besides these two fire rates of the two climbing fiber signals, the cortex signals from the two cluster groups also arrived in the Pontocerebellum (omitted in the figure above!). The cortex signals ran - topologically well sorted - over the bridge cores of the crossing floor to the moss fibers of the Pontocerebellum. These excited the cerebellar nuclei via collaterals and ended at the granule cells of the pontocerebellum. Each cerebellum cluster received its own climbing fibre, its Purkinje cells received the moss fibre input from exactly one cortex cluster.
Cluster theorem of the cerebellum
The signal neurons of a cortex cluster end in the associated cerebellum cluster at the granule cells of this cluster. Neighbourhood relationships between the clusters are largely retained. Each cerebellar cluster is supplied by a climbing fibre, which is derived from the mean signal of the cluster group by signal divergence. This cluster group signal of the nucleus subthalamicus is transformed by the striosome system of the basal ganglia into a clocked tetanic oscillation, which is distributed to the cerebellum clusters by linear signal divergence in the nucleus olivaris. In its cerebellum cluster, each climbing fiber contacts a series of Purkinj groups arranged one after the other, since it exhibits sequential divergence.
For example, if the degree of divergence in the nucleus olivaris is equal to four, four new climbing fibre signals are generated from each cluster group signal. Each of them arrives in one of the four cerebellum clusters. To each cerebellum cluster now belongs a series of Purkinj groups arranged one behind the other.
In humans, about 430,000 neurons of the subthalamic nucleus supply 15,000,000 Purkinje cells. Thus, there are about 36 Purkinje cells for every subthalamic neuron. If we divide these into four strips, each strip contains 9 Purkinje cells. If now three Purkinje cells form one Purkinje group, then one climbing fiber supplies 3 Purkinje groups. Since each Purkinje group can learn a certain complex signal, the Cerebellum would be able to learn about 5,000,000 such complex signals, or 5 million of them. Of course, this is not quite true, since part of the cerebellum, as a vestibulo- and spinocerebellum, is only used for signal inversion.
How many cortex clusters could be considered in a complex signal. A parallel fibre in humans reaches about 460 Purkinje cells, i.e. about 150 Purkinje groups. If three Purkinje groups belong to each cortex cluster, 50 cortex clusters remain. Then one Purkinje cell of 50 cortex clusters could be reached via the parallel fibre signals. Added to this is the range of the basket cells. Depending on the literature, a human basket cell can reach between 8 and 80 Purkinje cells. Thus, one Purkinje cell can reach the input of 400 to 4000 cortex clusters. This corresponds to a cortical rectangle whose edge has a length of about 20 to 63 cortex clusters.
If we look at the primary visual cortex and imagine the orientation columns for it, which belong to an eye dominant column in the cortex cortex cortex, we could think of this square area divided into 400 subsquares, i.e. 400 cortex clusters. In each of these clusters there is then a mean neuron from which the climbing fibre signal is derived. In the associated 400 cerebellum clusters, any Purkinje cell could receive the complete input from the ocular dominant column, i.e. the 400 associated cortex clusters, via the parallel fibres and the basket cells. Each of them could learn an orientation direction by imprinting. This would enable the Pontocerebellum to achieve a directional resolution of more than half a degree, since 180 degrees represents a complete rotation. The longer the parallel fibres are and the more Purkinje cells are reached by a basket cell, the finer the resolution in the Pontocerebellum will be. It is therefore not surprising that these sizes increase with progressive evolution.
As described above, each Purkinj group in the Cerebellum cluster was able to learn incoming complex signals, even before the signal divergence in the nucleus olivaris. The algorithm was the same. However, due to the signal convergence of the mean signals in the subthalamic nucleus, the fire rate of the global mean signals and thus of the climbing fibre signals is significantly increased, making LTD and LTP much more effective.
At this point we must note the inhibiting projection of the cerebellar nucleus to the olive pit. Each glutamatergic output neuron of the nucleus dentatus activated (at least) one inhibitory GABAergic neuron, which realized the lateral neighbor inhibition. Whether this neuron simultaneously projected inhibitively into the nucleus olivaris or whether a separate, separate projection neuron was activated with the transmitter GABA to carry out this inhibition in the olive ultimately makes no difference. In the case of recognition, the active output neuron inhibited the signal sources in the nucleus olivaris, i.e. the two signal-producing neurons that received the excitations fA and fB and divergently distributed them to the output line of the nucleus olivaris via intermediate inhibition neurons.
Since this was true for each output neuron of the Cerebellum cluster, all inhibition neurons of the cluster projected inhibitively into exactly the two output neurons of the nucleus olivaris, which provided all climbing fiber signals of the Cerebellum cluster via signal divergence.
This inhibition was not total, but relative. The previous tetanic excitation was weakened in its rate of fire to such an extent that it became low frequency and was no longer sufficient for further long-term depression or long-term potentiation in the Pontocerebellum. A total inhibition would be unfavourable, because permanently unexcited neurons would possibly be doomed. However, the remaining low-frequency residual excitation could lead to further interesting results due to the phenomenon of long-term potentiation (LTP). There are such effects that are triggered by low-frequency stimulation.
Theorem of inhibition of the nucleus olivaris by the nucleus dentatus
In the cerebellar nuclei, inhibitory interneurons increased the contrast of the output. In the nucleus dentatus, lateral inhibition was perfected in such a way that the master excited neuron of one neuron row completely inhibited its neighboring neurons in the same row. In addition, it excited an inhibitory projection neuron, which projected to the two input neurons of the nucleus olivaris to inhibit their mean excitation to such an extent that they did not exhibit tetanic excitation, but only low frequency excitation.
All output neurons of a neuron row were connected to this inhibitory output neuron, so that now, no matter which of them was active, the tetanic excitation of the corresponding two input neurons in the nucleus olivaris was reduced to a low-frequency level. This prevented a certain complex signal in the cluster from imprinting several Purkinj groups, i.e. multiple imprinting occurred
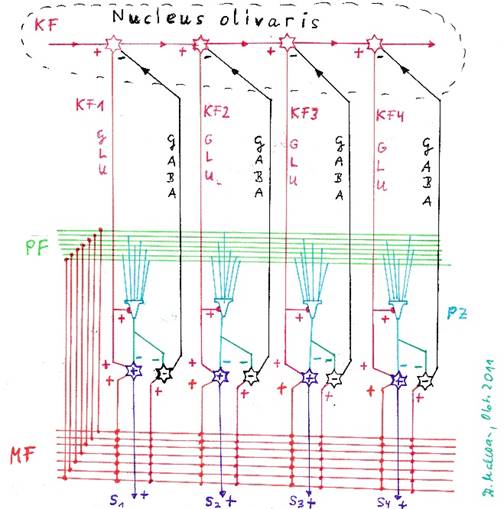
Abbildung 39 - Die Hemmung der Olive durch die Neuronen des Nucleus dentatus
Now we should define the terms embossing signal, intrinsic signal and external signal.
Definition: embossing signal, own signal, external signal
The cortical signal, which in a Purkinje group caused imprinting via LTD and LTP and led to the associated output neuron of the nucleus dentatus being the only one to react with a strong output, is called imprinting signal, the process is called imprinting.
Through the embossing process, the embossing signal becomes the signal of the Purkinj group concerned.
Any signal different from this signal is called an external signal of the Purkinje group.
The learning of new imprinting signals is a process in which several Purkinj groups are always involved and receive the same input. These groups are in competition with each other. The Purkinje group in which the processes of LTD and LTP cause the strongest synaptic changes emerges as the winner. The competitive situation is supported by several algorithms.
On the one hand, the climbing fibre signals with their tetanic excitation act on the golgi cells and excite them strongly. These golgi cellsinterrupt the signal flow to the granule cells, so that subsequently arranged Purkinje cells receive less input and their imprinting must be less. Thus, within a series of Purkinje cells it is ensured that the first unembossed Purkinje group is the first to reach the embossing threshold and interrupts the climbing fibre signal with its recognition signal. But the subsequent Purkinje cells have experienced incomplete imprinting. This can be reversed by further algorithms over time. The process required for this will be referred to here as de-embossing or embossing reversal.
The phenomenon of imprint reversal can occur in the cerebellum when the Purkinje cells are excited at low frequency and the parallel fibres are active at the same time. As a result, an increase in synaptic coupling between the active parallel fibres and the Purkinje cell occurs. However, this requires that the Purkinje cell is activated much longer or more frequently. Thus, low-frequency activation of a Purkinje cell and simultaneous activation of a parallel fiber for at least 300 seconds is required, according to recent research [117] by Varda Lev-Ram, Scott T. Wong, Daniel R. Storm and Roger Y. Tsien in 2002.
We now assume that the cortical imprinting signal may be active for a total duration of 300 seconds, with possible pauses in between.
Thus, the incomplete imprinting of the synapses between the parallel fibres and the Purkinje cells is reversed by increasing the degree of coupling reduced by LTD again by LTP.
For the synapses between basket cells and Purkinje cells or star cells and Purkinje cells, whose coupling degree had been increased by incomplete LTP, there is an analogous effect of long-term depression due to low-frequency climbing fibre exposure.
The incomplete inhibition of the climbing fiber signals when recognizing the intrinsic signal of a Purkinje group serves to reverse the incomplete imprinting in the neighboring groups in the Cerebellum cluster.
Theorem of the recognizing Purkinje cell
If a Purkinje group in the Pontocerebellum recognizes its cortical imprinting signal, its assigned dentate neuron immediately inhibits the climbing fiber signal arriving with a time delay via an intermediate inhibition neuron, so that it becomes low-frequency and is no longer available for renewed imprinting in the Pontocerebellum. The now low-frequency climbing fibre signal causes a (slow) de-embossing of the current embossing signal in the neighbouring Purkinje cells that have been incompletely embossed, by increasing the degree of coupling between the active parallel fibres and the Purkinje cells by LTP up to the value 1. For the synapses of the basket and star cells with the corresponding Purkinje cells, the degree of coupling is decreased to the initial value before partial imprinting. In the end, only one Purkinje group will react to this imprinting signal.
But what happens if it is not the embossing signal but a different signal that affects the embossed Purkinje cell?
Essential for the phenomena LTD and LTP is the time lead with which signal class 5 arrives in the Pontocerebellum before signal class 6. While the cortical signals of class 5 arrive at the granule cells via the crossing level and the moss fibres and activate the parallel fibres, the mean signals of class 6 have to take the much longer route via the substantia nigra pars compacta, the striatum, the nucleus ruber and the nucleus olivaris. Therefore they arrive with a significant time delay.
If a signal different from the imprinting signal arrives, this Purkinje cell is strongly excited by the incoming cortex signals, because the synaptic coupling between the parallel fibers and the Purkinje cell has the value 1. This strong excitation is built up before the climbing fiber signal can arrive via the tractus tegmentalis centralis. The strong excitation of the Purkinje cell completely inhibits the corresponding output neuron in the nucleus dentatus. There is no output as long as the climbing fibre signal has its blanking pause.
If the climbing fiber signal now arrives with a time delay, it is basically ineffective. The strong inhibition caused by the Purkinje cell causes a short circuit between the exterior and interior of the cell in the nucleus dentatus, the inhibition prevails against the excitation. Therefore, the output neuron does not provide any output. This applies to all Purkinje cells of the respective Purkinje group.
Theorem of reaction to an external signal
A imprinted Purkinj group reacts to the arrival of a foreign signal with a strong inhibition of the corresponding output neuron in the nucleus dentatus. This causes a short circuit in the output neuron, so that the delayed arrival of the climbing fiber excitation remains ineffective and no output occurs.
If, however, a complex signal consists partly of the intrinsic signal of a Purkinje cell and still contains parts of an external signal, the Purkinje cell is less excited by the intrinsic signal part than before imprinting, the inhibition of the output neuron is reduced and there is an output. However, this output is lower than if only the own signal had been present. This is a typical behavior of neural networks.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan