Theory of the brain’s neural circuitry
and analytical thinking
ISBN 978-3-00-037458-6 - ISBN 978-3-00-042153-2
Chapter 2: The multilevel parvocellular system of the brain
Dedicated in honourable memory of Professor Rolf Kötter, whose
extraordinary research achievement and his excellent descriptions of cytoarchitectonics
of the basal ganglia system provided the author with the inspiration and factual knowledge to understand how the basal ganglia function from an information theory perspective
Part 2.1 The parvocellular climbing fibre system of the brain
In chapter 1 the magnocellular mean value system of the brain was presented. It could be shown that the mean value signals of the cortex clusters are used to generate climbing fiber signals. These climbing fibre signals represent the neuronal write command for the Purkinje cells of the cerebral bellum, which leads to the storage of the current imprinting signal. By this write command the imprinting signal of the cortex becomes the intrinsic signal of the Purkinje cell. The intrinsic signals form the column vectors of the associative matrix.
The input vector of the cerebellum cluster consists of the signals of the signal neurons of the associated cortex cluster, which enter via the bridge nuclei. The output of the cerebellum cluster is a vector that results from multiplying the input vector with the associative matrix. Its individual vector components are formed by the output neurons of the cerebellum nuclei. These are active precisely when the average strength of the own signal of the associated Purkinje cell reaches at least the average signal strength of the current foreign signal. This is made possible by the interaction of the Purkinje cell with the associated basket and star cell as well as by the formation of single signals in the output neurons. The special feature of the cerebellar system is its imprinting ability. The climbing fiber signal derived from the activity neurons of the cortex cluster "burns" the imprinting signals into the Purkinje cells, basket cells, star cells, golgi cells and nuclear neurons. As a result, this signal becomes the intrinsic signal of the basket cells, star cells, golgi cells and nuclear neurons, but an external signal for the Purkinje cells. While the foreign signals excite the Purkinje cells, the own signals excite the basket and star cells. The latter inhibit the Purkinje cells. Depending on which type of signal predominates, the output neurons react differently. Therefore, the cerebellum can be interpreted as a neural associative memory with a special write command.
Since it can be shown that there are other types of climbing fiber signals in addition to the climbing fiber signals derived from the magnocellular mean value system, we need to specify the different types of climbing fiber more precisely.
Definition 2.1: Climbing fibre signals of the different systems
The climbing fibers derived from the output of the mean neurons (activity neurons) of the cortex cluster are called climbing fibers of the mean system, climbing fibers of the magnocellular system or optionally climbing fibers of the striosome system.
The climbing fibers, which are derived from the output of the signal neurons of the cortex, are called climbing fibers of the parvocellular system or alternatively climbing fibers of the matrix system.
The climbing fibers, which are derived from the output of the cerebellar nuclei, are called climbing fibers of the cerebellar system.
The climbing fibres that project directly from the receptor systems via the olive into the cerebellum are called climbing fibres of the spinal system or climbing fibres of the receptor system.
In this chapter we analyze the climbing fiber system of the parvocellular system of cortex clusters.
But before the author introduces the parvocellular climbing fibre system, which primarily serves to generate short-term memory and is one of the main tasks of the basal ganglia, all neurologists should have the opportunity to get to know the cytoarchitectonics of the basal ganglia from the author's perspective.
The author owes most of his insights on this subject to Professor Karl Zilles on the one hand and Professor Rolf Kötter on the other. The latter was a gifted student of Karl Zilles and left us with an incredibly exact and inspiring description of the cytoarchitectonics of the basal ganglia.
Without his detailed descriptions of the individual substructures and their exact cytoarchitectonics, this monograph would never have been written.
The starting point is a graphic representation of the basal ganglia system according to Young and Penney from 2000, which was found in the very readable dissertation by Inka Eisfeld. What is special about this representation is that the nucleus pedunculopontinus with its acetylcholinergic projections was also included and all important projection paths are included, which are often missing in other representations.
The reader should understand the reproduction of the representation as a "pictorial quotation", which should be possible in the context of a scientific work even without the explicit consent of the authors.
Picture quote: Basal ganglia scheme according to Young and Penney:
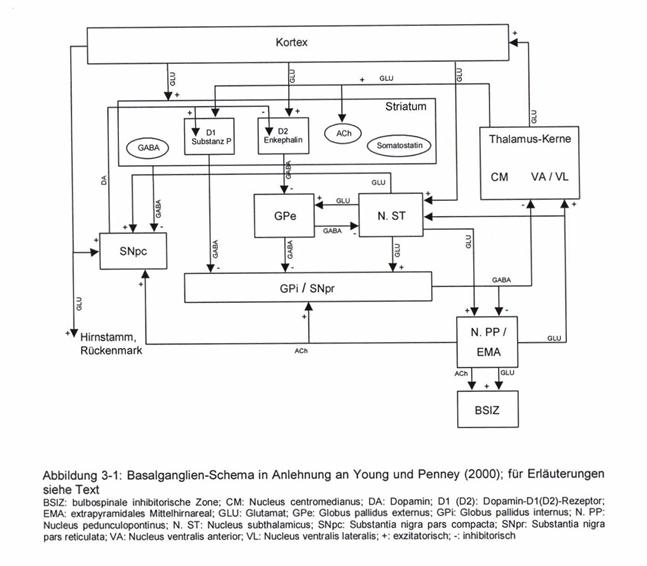
This presentation was the starting point of a scientific debate, as a result of which the author divided the basal ganglia system into two subsystems:
- the magnocellular striosome system of the basal ganglia
-
the parvocelular matrix system
of the basal ganglia.
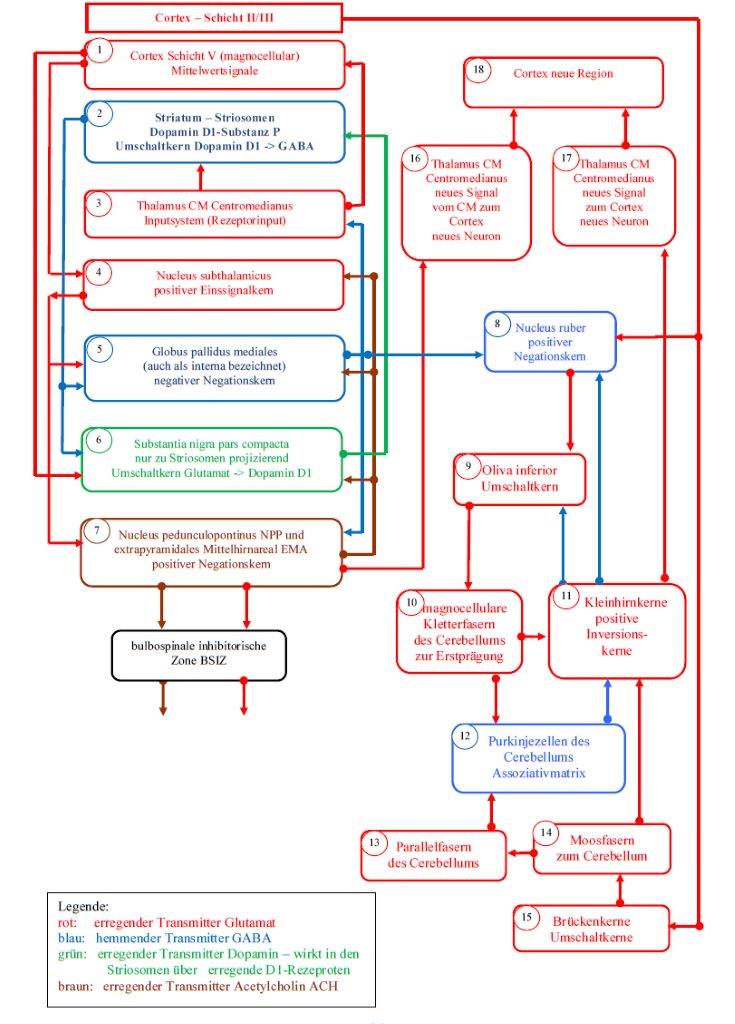
Sketch 2.1: Basal ganglia model of the striosome subsystem according to Andreas Heinrich Malczan
Sketch 2.2: Basal ganglia model of the Matrix subsystem according to Andreas Heinrich Malczan
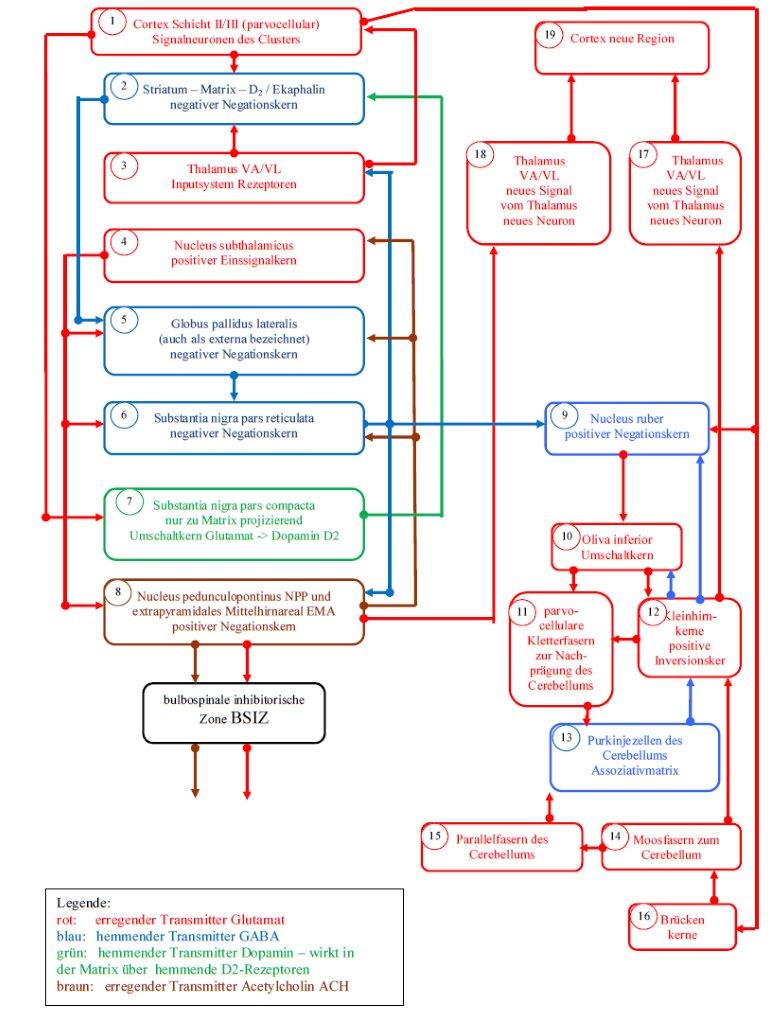
The representations of the striosome system and the matrix system are organized as flowcharts. To facilitate explanation, the sub-elements are labeled and marked with small numbers, which are located within the small circles. The different colours represent the different transmitters, which are specified in the corresponding legend.
The proof that the structural and system-theoretical connections presented by the author are close to reality primarily uses the insights presented by Professor Rolf Kötter.
At this point it is worth noting once again that the author of this monograph is a mathematician. In his theory, he applies the findings that thousands of neurologists have worked hard to develop themselves.
The following statements in this section are largely based on a script by Prof. Dr. Rolf Kötter, which is available at
https:/www.hirn.uni-duesseldorf.de/~rk/Taeching/lect_bg.htm
respectively under
https://www.anatomie.net/Unterricht/Skripte/lect_bg.htm
are available (in July 2011).
But first, we recapitulate the theorem 1.2 of this monograph and reproduce it again (as theorem 2.1):
Theorem 2.1: The substantia nigra pars compacta is a switching core for the cortex output
The substantia nigra pars compacta receives on the one hand the output of the signal neurons of the cortex cluster and on the other hand the output of the activity neurons. Both types of out-put are glutamatergic and are switched to the transmitter dopamine. While the cluster neurons are switched to dopamine of the subtype D2 and inhibit their target neurons in the matrix of the striatum, the activity neurons of the cortex clusters are switched to dopamine of the subtype D1 and excite their target neurons in the striososomes of the striatum.
With regard to the matrix system, theorem 2.1 claims that the signal neurons of each cortex cluster move to the substantia nigra, where they switch to dopamine of subtype D2. In this process, the axons of the signalling neurons traverse the neuronal core area of the striatum, so that their excitation is available there.
The axons of the substantia nigra pars compacta move back towards the cortex, but end before that in the core area of the striatum. This consists of two subsystems that can be easily separated spatially. One subsystem is the striosomes, the other the matrix.
Here one should imagine that the striosomes represent small spatial areas embedded in the matrix, similar to raisins in a cake. Both subsystems are distinguished by the receptors of their synapses. In the striosomes, the dopamine has a stimulating effect via D1 receptors. In the matrix, however, it has an inhibitory effect via the D2 receptors.
The inhibited neurons of the striatum are rightly referred to in the literature as main neurons. In the work "Functional Neuroanatomy" by Karl Zilles and Gerd Rehkämper we read on page 278 of the third edition:
(begin quote)
"The striatal target cells of the corticostriatal tract are medium-sized neurons (soma diameter 15 µm), which make up more than 70 % of the nerve cells in the corpus striatum and are therefore called main neurons. Their dendrites are densely packed with spines and each main neuron receives thousands of different cortical axons. Main neurons usually show little electrical activity because they only form action potentials when many monkey borders on their dendrites are active at about the same time. However, when they generate action potentials, they have an inhibitory effect on their target neurons by releasing the transmitter GABA.
(end of quote)
We remember the definition 1.14 of the negative negation nucleus with internal single signal formation. Therefore we make the following assumptions:
- The main neurons of the matrix of the striatum are single signal neurons. They tap into the passing and exciting cortex output, forming huge dendrite trees. A possible pure averaging can be excluded, since we are talking explicitly about thousands of excitatory axons passing by. The single signal is negative because the main neurons are GABAerg.
- The main neurons thus form a negative input signal, which is inhibited by the axons of the nigrostriatal input. This is a typical characteristic of nerve-ion nuclei.
- Any D2-dopaminergic signal from the substantia nigra pars compacta is therefore negated.
- Since the output of the neurons of the substantia nigra pars compacta of subtype D2 is switched to dopamine by the signal neurons of the cortex, the cortex output of the signal neurons in the matrix of the striatum is ultimately negated.
Therefore the following knowledge of the author applies.
Theorem 2.2: The matrix of the striatum is a negative negation nucleus with internal single signal formation
The matrix of the striatum is a negative negation nucleus for the output of the signal neuroses of the cortex clusters. Coming from the cortex, this is first conducted to the substantia nigra pars compacta and there it is switched to dopamine of the subtype D2. After switching over, it inhibits the main signal-forming neurons of the matrix, whereby this cortex output is negated. Since the main neurons transmit the negated signal via GABA, the matrix is a negative negation nucleus. An internal single signal formation is present because the single signal is derived from the traversing cortex output.
Later it will be shown that the proven connection between matrix and strioso-men by the transmitter acetylcholine (ACH) has an important role in attention control.
In sketch 2.2 you can see that the output of the matrix (2) first moves to the globus pallidus lateralis (5), which is also called pallidus externa in the literature. However, this core area receives input from the subthalamic nucleus (4). It was shown that the subthalamic nucleus is a positive single-signal nucleus according to theorem 1.4.
A region that receives excitatory input and inhibitory input on the one hand and produces only inhibitory output on the other hand, can in all probability only be a negative negation nucleus. Another possibility would be an inversion kernel.
But as the output of the Globus pallidus externa is not permanently active - which would be true for an inversion kernel - only the negation kernel remains. Thus we formulate a new theorem.
Theorem 2.3: The globus pallidus externa is a negative negation nucleus for the GABAergic output of the matrix of the striatum
The globus pallidus externa is a negative negation nucleus with external single signal formation. The positive single signal originates from the nucleus subthalamicus. The output of the Globus pallidus externa is the double negated output of the signal neurons of the cortex clusters. Since the output is inhibitory, there is a negative negation nucleus.
After the output of the signal neurons of a cluster was negated for the first time in the matrix of the striatum, it is now negated again in the globus pallidus externa. On the one hand, this results in a time shift of the cortex signals, since the action potentials have to cover a considerable distance. On the other hand, a frequency shift occurs, whereby the original firing rate of the cortex neurons is raised to a medium signal level or even lowered. The latter is caused by the generated single signals.
In sketch 2.2 it can be seen that the output of the globus pallidus externa (5) is directed to the substantia nigra pars reticulata (6). However, the latter in turn receives the input signal from the subthalamic nucleus (4).
With the same argumentation it follows that the substantia nigra pars reticulata is also a negative negation nucleus with external input signal.
The output of this nucleus moves to the nucleus ruber (9). This nucleus has already been shown to be a positive negation nucleus. Therefore the following two theorems apply:
Theorem 2.4: The substantia nigra pars reticulata is a negative negation nucleus for the GABAergic output of the globus pallidus externa
The substantia nigra pars reticulata is a negative negation nucleus with external one signal formation. The positive one signal originates from the nucleus subthalamicus. The output of the substantia nigra pars reticulata is the triple negated output of the signal neurons of the cortex clusters. Since the output is inhibitory, there is a negative negation nucleus.
Theorem 2.5: The nucleus ruber is a positive negation nucleus for the GABAergic output of the substantia nigra pars reticulata
The nucleus ruber is a positive negation nucleus with internal single signal formation. The positive single signal is formed in the nucleus ruber itself. For this purpose, the magnocellular output of the mean value neurons (activity neurons) of the cortex cluster, which originates from the cortex, is used to create the magnocellular part of the nucleus ruber. The positive single signal is inhibited by the output of the substantia nigra pars reticulata and produces as output the negation of the signals from the substantia nigra pars reticulata. Since the output is excitatory, a positive negation nucleus is present.
Thus, the output of each signal neuron of the cortex cluster is negated four times in total:
- for the first time in the matrix of the striatum (negative negation nucleus)
- for the second time in the globus pallidus externa (negative negation nucleus)
- thirdly, in the substantia nigra pars reticulata (negative negation nucleus)
- fourth, in the nucleus ruber (positive negation nucleus).
From the nucleus ruber, the signal-carrying axons move to the inferior oliva, release collateral axons to the cerebellar nuclei concerned as usual and reach the Purkinje cells as climbing fibres.
Therefore the following knowledge applies:
Theorem 2.6: The output of the signal neurons of a cortex cluster reaches the Purkinje cells of the associated cerebellum cluster fourfold negated, time-delayed and frequency-transformed as climbing fiber signal
The signals of the signal neurons of a cortex cluster are quadruple negated by the matrix system of the basal ganglia and transformed into climbing fiber signals, whereby they are delayed by a considerable time difference.
The cerebellum thus possesses two variants of each imprintable signal of the cortex cortex of a cortex cluster:
- a moss fibre variant, which could be described as a direct variant, in which the cortex signals reach the moss fibres with a very short time delay via the bridge nuclei, which both supply the neurons of the cerebellum nuclei and excite the parallel fibres via the granule cells
- a climbing fibre variant which could be described as an indirect variant Here, the cortex signals reach the substantia nigra pars compacta, the matrix of the striatum, the globus pallidus externa, the substantia nigra pars reticulata, the nucleus ruber and the nucleus olivaris inferior with a greater time delay, and finally the cerebellar nuclei and the climbing fibres of the matrix system.
Now it is time to estimate the magnitude of the time delay imposed on the cortex signals by the matrix system. Here we recall the theorem 1.13. Among other things, it stated that the system clock in the striosome system is about 5 Hz. This finding is supported by the statements of Karl Zilles and Gerd Rehkämper about the loose coupling of the neurons of the substantia nigra pars compacta by dendrodendritic synapses.
In the figures on page 31, the "slow-motion display" shows that the action potential requires 100 ms for a complete loop run, from which the cycle duration of 200 ms and a cycle frequency of 5 Hz are derived.
If a complete loop takes 100 ms, a signal from the cortex to the substantia nigra pars compacta takes about 50 ms, from there again 50 ms to the striatum, and possibly an estimated 25 ms for the quadruple negation.
Thus 125 ms would be sufficient for the distance from the cortex to the Purkinje cell. This would correspond to the value of 1/8 second.
All neurologists are free to correct this figure estimated by the author by exact measurements. We summarize this estimated result in a separate theorem.
Theorem 2.7: Time delay of cortex signals by the matrix system
The time delay of the signals of the signal neurons of the cortex through the matrix system of the basal ganglia on their way to the Purkinje cells of the cerebellum is estimated to be about 125 milliseconds. Only after this time delay does the signal of a signal neuron from the associated cortex cluster arrive at the associated Purkinje cell as a climbing fibre signal.
In sketch 2.1, but also in sketch 2.2, there is still the neuron nucleus called nucleus pedunculopontinus (NNP), whose tasks have not yet been addressed. This must now be made up for.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan