Theory of the brain’s neural circuitry
and analytical thinking
ISBN 978-3-00-037458-6 - ISBN 978-3-00-042153-2
Part 2.2 The role of the nucleus pedunculopontinus in the basal ganglion system
In both sketch 2.1 and sketch 2.2 for the basal ganglia model, the nucleus pedun-culopontinus is part of the system. This already applies to the basal ganglia schema by Young and Penney from the year 2000.
A closer analysis and comparative considerations led the author to the similarity between nucleus ruber and nucleus pedunculopontinus:
- Both cores use excitation input signals. The nucleus ruber forms these itself from the input of the mean value neurons of the many cortex clusters, the corresponding single signal neurons form its magnocellular part. The nucleus pedunculopontinus receives its external input signal from the nucleus subthalamicus.
- Both nuclei receive inhibitory input from the globus pallidus interna of the striosome system (see sketch 2.1).
- Both nuclei receive inhibitory input from the globus pallidus externa of the matrix system (see sketch 2.2).
- Both nuclei produce excitatory output, but the output of the nucleus ruber is glutamaterg, while the output of the nucleus pedunculopontinus uses the excitatory transmitter acetylcholine.
The excitatory signals of the nucleus ruber reach the Purkinje cells via the olive as climbing fibres. In contrast, the acetylcholinergic output of the nucleus pedunculopontinus reaches the nucleus centromedianus of the thalamus in the striosome system (sketch 2.1). In the matrix system, this output also reaches the thalamus, but this time the thalamus VA/VL.
The abbreviation VA stands for Nucleus ventralis anterior, VL stands for Nucleus ventralis lateralis. Both are thalamic nuclei.
As is well known, almost all neurologists hold the doctrine that the thalamus is the gateway to consciousness, whose headquarters are located in the cortex cortex. In this respect, the author is not surprised that the laboriously produced output in the basal ganglia system also reaches the thalamus as an excitatory signal. However, the striosome system and the matrix system also have different target partners in the thalamus. Therefore a certain division of tasks is to be assumed.
At thispoint, it is worth recalling the dedication in Chapter 1. It refers to a thought model by Prof. Ulrich Ramacher. He designed the model of a dissolution pyramid. How should one imagine this?
The starting point for an explanation could be a simple picture. So we think of 1000 pixels in a row next to each other and 1000 such rows of pixels below each other. Therefore the image has a resolution of 1,000,000 pixels, that is one million pixels.
If such an image were converted one-to-one into neuron signals, one million receptors with associated neurons would be needed. We want to call these neurons start neurons.
Now the mean value neurons come into play. Let us imagine that a mean value neuron would tap 25 of the start neurons and calculate a weighted mean value from them. We call these mean value neurons the first stage mean value neurons. Their receptive field is 25 pixels, that is 5 pixels wide and 5 pixels high.
However, this divides the original image width of 1000 pixels by 5, so that we now find 200 first-stage mean value neurons in the image width.
Likewise, we will find only 200 average values in the image height instead of 1000 pixels.
If we now use the mean value neurons instead of the start neurons, the original image with 1,000,000 pixels corresponds to a first-level mean value image with 40,000 pixels. This image therefore has a lower resolution. If we sent it as input to the thalamus centro-medianus like the original image, we would have two images of the visual world: one in original resolution and one "coarser", i.e. also "reduced", image. Thus, each mean neuron not only determines the mean activity in the corresponding cluster, but is an output deliverable for a compressed image in the next level of an imaging pyramid. It should not be forgotten that the cortex clusters may very well overlap partially.
Sketch 2.3: Dissolution pyramid according to Ramacher as picture quotation:
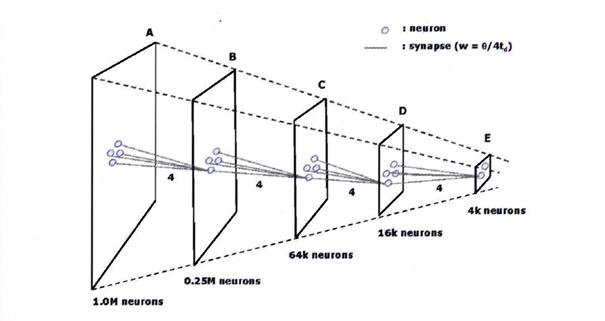
Source: "On the construction of artificial brains", Ulrich Ramacher / Christoph von der Malsburg, 2009, Springer, page 323, Figure 16.5
Are such reduced images now fantasies of the author, pure speculation or reality?
Such a phenomenon has been observed in the thalamus and cortex cortex. This is what Zilles/Rehkämper write in the "Functional Neuroanatomy" by Springer-Verlag on page 230:
(begin quote)
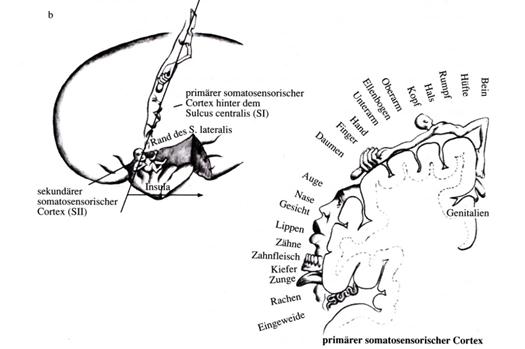
"The secondary somatosensory cortex S II contains the representation of the ipsa and contralateral sides of the body in low spatial resolution. “
"The small, secondary field SII also shows a somatotopic representation of the periphery, albeit with less spatial resolution and based on a less differentiated architectural structure. The connections are basically similar to those in S I, except that here a representation of both halves of the body is found, not only the contralateral side as in S I. “
(end of quote)
The author would like to supplement this quotation from "Functional Neuroanatomy" by Zilles/Rehkämper with a subsequent pictorial quotation from the extremely readable work "The Brain" by Richard F. Thompson of Spektrum Akademischer Verlag. It is taken from Figure 8.16 on page 268. We would like to take this opportunity to thank the publisher for the excellent translation of the original work from English. Without this translation the author would never have dealt with neurology, because this work was the first neurological textbook he read. Had it only been available in English, this manuscript would not have been written.
Sketch 2.4: Picture quote from Thompson: Representation of the field S I and the corresponding, reduced field S II in the somatosensory cortex of humans
The author considers it important to note that the process of "scaled-down mapping" is recursive, i.e. it is repeated continuously until only an average value remains from the original image. This is based on the assumption that the cortex cortex has a similar basic structure everywhere. So if the nucleus centromedianus thalami sends the mean value signals of the reduced image to a new cortex area, cortex clusters will also be present there. These supply input to new mean value neurons of layer V, which in turn return to the nucleus centromedianus via the basal ganglia system. In this way, everything repeats itself until finally only one mean value neuron remains.
We summarize our findings in a new theorem. As holder of the original rights to this knowledge, the author names this theorem after the inspirer, Prof. Ulrich Ramacher.
Theorem 2.8: Theorem of Ramacher
The projection of the nucleus pedunculopontinus to the nucleus centromedianus is part of a recursive system of a resolution pyramid, which recursively produces new, reduced images with lower resolution from the original image. The basis for this is formed by the mean neurons of layer V of the cortex cortex, which via the striosome system ultimately send the input for a coarser image back to the thalamus centromedianus, from where it reaches the cortex.
This would clarify the exciting projection of the striosome system (sketch 2.3) from the nucleus pedunculopontinus (7) to the thalamus centromedianus (16) and then to the cortex.
However, the exciting projection of the matrix system (sketch 2.2) from the nucleus pedunculopontinus (7) to the thalamus VA/VL and from there to the cortex remains open. Which signals are sent by the NPP to the cortex via the thalamus.
Here we remember the similarity between the nucleus ruber and the nucleus pedunculopontinus. According to theorem 2.5, the nucleus ruber sends the quadruple negated output of the signals of the signal neurons of the associated cortex cluster (1) to the olive, where they become climbing fiber signals.
The nucleus pedunculopontinus, just like the nucleus ruber, adopts the last of the four negations. For up to the substantia nigra pars reticulata (6) of sketch 2.2, the primal signals of the cortex were negated three times in total. Now they are negated a fourth time in the nucleus pedunculoponti-nus and are transmitted as an excitatory signal to the thalamus VA/VL, from where they reach a new cortex region.
Thus, not only the cerebellum receives the quadruple negated original signal of the cortical signal neurons, but also the cortex (in a new projection area).
We remind you that the quadruple negated signal is now strongly time delayed and frequency transformed. So the cortex also receives an image of all "signals of the past", which is about 125 milliseconds ago.
Theorem 2.9: Time delayed projection of the signals of the signal neurons of a cortex cluster into a new cortex region
The projection of the nucleus pedunculopontinus via the thalamus VA/VL to a new cortex cluster is organized in such a way that the output of each signal neuron of the original cluster reaches the new cluster as input, whereby the signal is fourfold negated, time-delayed and frequency-transformed in the matrix system. The time delay has approximately the same order of magnitude as the time delay of the same signals on their way to the cerebellum via the nucleus ruber and the olive, but the latter signals are the climbing fiber signals of the parvocellular system.
Part 3 and part 4 of this monograph show how the output of the NPP is used to detect and digitize temporal changes of the signal position, mainly of movements of visual and also motor objects (e.g. joints), so that they can be stored and retrieved in the cerebellum. Then it will also be explained what the task of the millions of connections between the cortex areas is. Provided that the associated theory proves to be faultless on examination.
Both the cortex and the cerebellum thus receive the signals of the signal neurons of the cortex, which are delayed by the matrix system. Since the original cortex signals are becoming increasingly important in systems theory, we want to give these signals another name that is relevant in systems theory.
Definition 2.2: Elementary signals and complex signals
The signal of a signal neuron of a cortex cluster is called elementary signal. Thus, each signal neuron Nk of the cluster corresponds to an elementary signal, which can be unambiguously described by specifying the firing rate fk and its Index k.
A signal vector S whose components contain the fire rates of all elementary signals of the cluster according to
S = ( f1, f2, f3, ..., fn),
represents a complex signal K of the cluster.
In general, a complex signal is regarded as a "snapshot" because the fire rates of its components can change over time.
Now we can make an analysis of the signals and propagation paths.Monografie von Dr. rer. nat. Andreas Heinrich Malczan