Vertebrate brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-064888-5
3.5 The topological well-being of neurons in the launch system
Over the course of evolution, animal organisms have produced an enormous variety of receptors with which the most diverse environmental stimuli can be transformed into neuronal signals.
If the number of different receptor types was initially small, and there were initially only a few receptors for each of them, a double rope ladder system in the form of a tetraneural nervous system was able to transport the signals quite well in the segmented bilateria. However, if the number of receptor types increased and the number of receptors also increased enormously, then a single neuron strand could not even absorb the huge number of axons. The strand, let's take the afferently projecting sensory strand of one side of the body, increasingly became a kind of flat ribbon cable of axonsarranged in a thick layer. The four nerve cords of the tetraneural system, two sensory and two motor, formed a kind of tube, the inside of which was without function and formed a cavity. This was filled with a liquid that was incompressible and could withstand the high water pressure even at greater depths. This is how the neural tubeof future vertebrates was formed.
Theorem of neural tube formation
The bilaterally symmetrical neural tube developed from the tetraneural nervous system by a very strong increase in both the number of receptors of one type of receptor and the number of different types of receptors in the course of evolution. The left and the right half of the neural tube each consists of the parallel connection of many individual rope ladder systems, the sensory and motor centers of the segments are interconnected in the manner described above. All structural elements of the tetraneural nervous system are realized analogously in the neural tube, such as the mean nuclei. Contralateral inhibition is also realized. The neural tube and the tetraneural nervous system of the segmented bilateria, whose line leads to the vertebrates, differ (initially) only in the much larger number of neurons. The reason for this is the enormous increase in the number of receptors of both the same modality and different modalities.
In the neural tube, the sensory connectors form the posterior horn, the motor connectors the anterior horn.
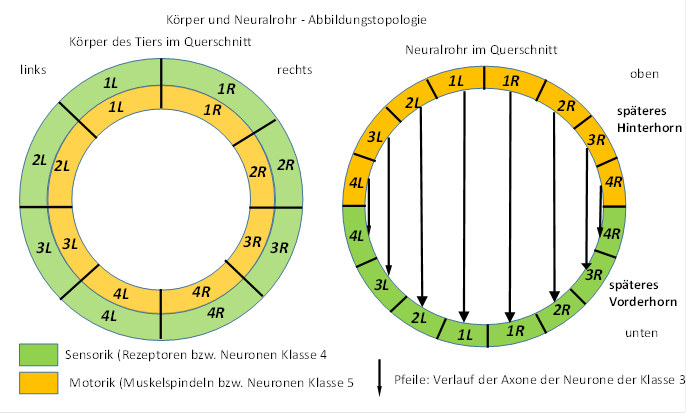
Figure
6 - Body and neural tube - imaging topology
directional theorem of the neural tube nervous system
In an idealized neural tube nervous system, the axons of the connective neurons form right angles with the axons of the commissure neurons. Spatial deformations of the body can lead to topological deviations from the ideal directions.
The results of the American Connectome project, in which the course of the axon in the human brain is investigated by computer tomography of the diffusion gradients of water molecules, confirm this spatial orientation. Axons of the same orientation run uniformly next to each other without mixing. Axons that branch off often form right angles to the previous axon course.
classification theorem for neurons
The directional theorem makes it possible to assign the projection neurons to the six different neuron classes solely on the basis of their axon course. Neurons with axons that run upside down belong to neuron class 4, while axons that run down belong to neuron class 5. Horizontal axons belong to neuron classes 3 and 2, where the axons of class 3 neurons target motor targets, while class 2 always project to the opposite side into analog neural centers. Axons of the class 6 neurons move to mean centers, while the neurons of the mean centers send out their axons to the class 1 neurons.
To a certain extent, axons of class 4 and 5 neurons could be confused with class 6 and 1 neurons if they were only viewed in small areas without knowing their termination neurons. This ambiguity disappears as soon as the source and target structures are recognized.
If the neural tube were cut along its longitudinal axis, the cut being slightly laterally oriented and cutting through both the posterior horn and the anterior hornon one side of the body, the sectional view would return to the original rope ladder system. Of course, the impression of the original segmentation islost due to the enormous number of neurons, since the neurons of a sensory or motor centre move up and down when there is a lack of space, where the neurons of the neighbouring segments are already trying to assert their place. This creates a surface on which the original sensory and motor centres are distributed.
However, the original segmentation can still be clearly seen in the spinal ganglia, which enter the neural tube as independent axon bundles.
We divide the neural tube into a trunk part and a head part. We now imagine any sectional plane that divides the body of the animal into two parts. The direction of the cut should be perpendicular to the body axis. In the sectional image - which looks roughly circular - we examine the arrangement of the axons of the upward and downward projecting connective tissue.
At first, we restrict ourselves to the axons of one modality. Their signals originate from a selected type of receptor, for example the muscle spindles. These receptors were formed relatively late; before that, free nerve endings took over the measurement of muscle tension.
Theorem of modality rings and segmental rings of axons in the trunk part of the neural tube
The axons of the upwardly projecting class 4 neurons and the axons of the downwardly projecting class 5 neurons of each modality form a modality ring in the neural tube. Within each modality ring there are as many segment rings as there are segments with the receptors of that modality.
The segment rings of a modality are nested and well-ordered. The segment ring with the highest segment number is located inside, whereby the numbering starts at the header segment. Towards the outside, the segment number decreases continuously. The outermost segment ring contains the axons whose receptors are located in the highest head segment.
In cross section the neural tube resembles a tree trunk with annual rings, but here groups of adjacent annual rings are combined to form modality rings.
The reason for this well-being in the neural tube is that the first type of receptor that was produced contacted the class 4 neurons for the head-side projection. Their axons simply grew straight up. Each higher segment attached its sensory projection axons externally to the existing ones. Thus each segment resulted in a segment ring of ascending axons. The motor part of the neural tube was analogous. Here the descending axons to the lowest tail segment were located inside and formed the innermost segment ring of this modality. The axons to the second lowest segment were located outside the described ones and formed the next segment ring. This continued until the first head segment was reached. The axons of his class 5 down-projecting neurons also attached themselves to the outside of the existing segment ring system and formed the outer segment ring. The entirety of all segment rings of the first modality thus formed a modality ring at the level of the first segment in the sectional image.
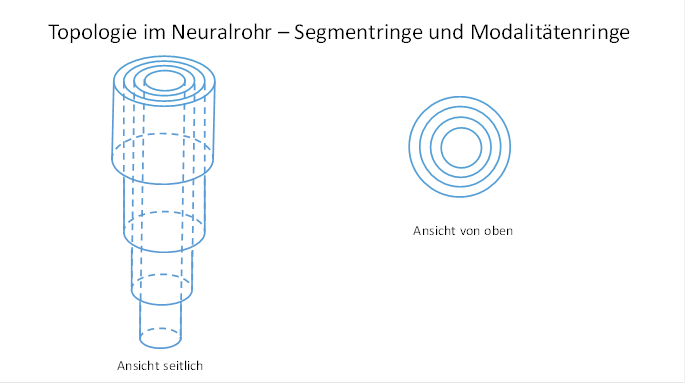
Figure 7 - Topology in the neural tube - segment rings and modality rings
When a second type of receptor formed, its projection axons were attached to the outside of the first modality ring. This in turn was done in the described way in segment rings that were coupled to the height of the segment.
The modality rings of the different modalities allow us to draw a conclusion about the order of their evolutionary formation. The modality rings of the earlier developed receptor types were on the inside, those of the evolutionary later developed receptor types on the outside.
Each segment ring consisted of four quarter rings, because in the original tetraneural nervous system there were two sensory and two motor axonal strands that formed the connective tissue and projected upwards and downwards respectively.
After we have described the axon course and the well-being order of the axons, we turn to the spatial arrangement of the involved neurons that possess these axons.
Theorem of segmented body images in the trunk area
In each segment, the sensory and motor connective neurons of the different modalities form an approximately cylindrical mantle surfacethat represents a topological body image of the signal-providing receptors of the segments. On the sensory side, a sensory body model is created on the outer surface of a half cylinder, while an analogous motor body model is created on the motor side.Neighborhood relationships of the receptors are preserved in the sensory body image. Neighborhood relationships of the target muscles are preserved in the motor body image.
We combine the sensory and the motor part of the body model of a modality to a modality cylinder, on whose mantle surface the sensory and motor body image is located. The modality cylinders of the different modalities are nested within each other, with the spinal canal of the neural tube running inside. Inside are the evolutionary older modalities, further outside are the evolutionary younger modalities. The parting plane between the sensory and the motor part of the neural tube is the plane of symmetry, each orthogonal straight line connects equal body points of the two images. All axons of the class 3 neurons are also orthogonal to this parting plane.
In each segment there is such a structure of nested modality cylinders. Since these receive their input from below (on the tail side), only the segments located below can be included in the body models. In contrast, the head segments contain a body image of all trunk segments for each modality. Only there a signal processing can include the signals of the whole body. Therefore the brain must be located on the head side.
For trunk modalities, only the signal-providing receptors are included in the sensory body image in the segment with the number k (counted from the head), i.e. only segments with the numbers k to n, if n represents the number of segments. The receptors located above the segment in question (normally) only supply the higher segments.
At this point it is appropriate to consider the topology of the nuclei of the seventh segment, which was the input and output segment of the primordial brain. It contained important projection nuclei, such as the nucleus ruber, the nucleus olivaris and the nucleus Purkinje, from which the later Cerebellum was to develop.
Theorem of motor body images in the nucleus ruber, nucleus olivaris and nucleus Purkinje
The motor connective neurons of class 5 of one half of the body form in the seventh segment the mantle surface of a half cylinder, which represents a topological body image of all ipsilateral motor neurons of the living being, which was ordered according to segment rings. Neighbourhood relations between the muscles and the motoneurons are preserved. In addition to this neuron surface, there is also a second mantle surface formed by the commissure neurons of class 2, which represent the beginning of the neuron chain for contralateral inhibition, which runs in a point-to-point connection via the nucleus olivaris to the purkinje nucleus and from there to the contralateral nucleus ruber.
The axons of the class 2 neuron layer of the nucleus ruber end in the nucleus olivaris.
Since they are unmixed, the neurons of the nucleus olivaris also form a body image of the motor neurons of the ipsilateral half of the body, well ordered by segment rings. The axons of the projection neurons transfer this topology to the Purkinje nucleus.
Initially, the Purkinje nucleus and the nucleus ruber are directly adjacent, later the neurons GABAergic neurons of the Purkinje nucleus separate and form an independent nucleus. The projection axons of his commissure neurons run horizontally well ordered to the nucleus ruber without crossing each other.
The Purkinje nucleus projected according to the same scheme in a point-to-point inhibitory image into the contralateral nucleus ruber, where it caused the contralateral inhibition of the associated motor opponents. Each GABAergic axon docks to the class 5 projection neuron in the nucleus ruber, which projects to the motor antagonist. Thus, contralateral inhibition is realized.
Due to the strong increase in the number of neurons involved, all of these one- or two-layer semi-cylinder surfaces, which actually represent neuron layers, were continuously deformed, compressed, and sometimes folded (to save space) in the course of further evolution. This gave them a functional appearance. However, the principle of imaging was retained in principle. Each core formed a well-ordered body image according to segment rings. The nucleus ruber, the nucleus olivaris and the nucleus Purkinje represented well-ordered body images of the motoneurons of one half of the body according to body segments, whereby a point-to-point connection was generally maintained.
Bilateralism was maintained here as well, with each of the cores being represented ipsi- and contralateral. Thus it was not impossible for the cross-commissures to cross each other in reality, even if the axons otherwise ran unmixed.
The arrangement in semi-cylindrical structures was favorable, so that there was enough space to let the horizontally running projection axons run from one nucleus to the next in the direction of the contralateral nucleus ruber without crossing and without mixing until the target nucleus on the opposite side was reached, whereby they naturally had to meet the axons of the opposite side. The involved axons of the class 2 commissure neurons ran undisturbed horizontally, while the axons of the class 4 and 5 projection neurons ran vertically upwards and downwards, respectively, without interfering with each other (when the head was up).
When considering the topological well-being in the starting system, neuron classes 1 and 6 were deliberately omitted in order not to unnecessarily complicate the model. In retrospect, it is not difficult to insert these neuron classes into the model, as well as the inhibiting interneurons that served as lateral neighbor inhibition.
In the course of evolution, other signals were to find their way into the olive pit, some of which were non-motor signals. The point-to-point connection was only present at the beginning and was lost at the latest in the second expansion phase of the developing cerebellum.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan