Vertebrate brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-064888-5
3.4 The basic structures in the primordial brain of the segmented Bilateria
The uppermost commissure, which connects the sensory center with the motor center in the head segment, will be called the cortical turning floor. We consider the sensory and motor center of the head segment as part of the cortical turning floor. In a longer developmental process, the vertebrate cortex emerged from it, which in turn can be divided into a motor and a sensory cortex, whereby the sensory cortex is well-ordered according to modalities. This is demonstrated in the following chapters. The mean core of the olfactory cortical floor is the septum.
The analogous structure, which already emerged from the fusion of the head-side segments, is called the thalamic turning floor. From it the thalamic system of vertebrates developed. The mean centre of this floor, which also exists bilaterally, is formed by a mean nucleus, which is called nucleus subthalamicus in vertebrates. We will already use this name here.
The sensory and motor centre on the third floor, together with the associated commissures, form a structure that we call the optical tectum. It represents (counted from above) the third turning floor. In it, there is signal interaction between motor and visual signals.
We assign a structure called the torus semicircularis to the fourth turn. Here the signals of the vestibular sense and (if present) the lateral line system act on the motor signals.
We reserve the fifth turning floor for a structure which will be described later and which we will call the crossing floor.
We reserve the
sixth
turning floorfor another pair of eyes. Here we postulate
that no lens eyes are used here, but convex facetted eyes.
In this floor we also place the optical mean value system, which supplies the
nucleus suprachiasmaticuswith signals.
The seventh turning floor(from above) represents the entrance systemor the entrance floor. Its sensory centre represents the input core of the primordial brain and receives the receptor signals of the trunk segments . In vertebrates, this input nucleus is split into two sub-nuclei, called nucleus cuneatusand nucleus gracilis. The former receives the receptor signals of the upper half of the trunk, the other receives those of the lower half. At the same time, this segment on the motor side is the initial level.
The motor centre of the seventh segment represents the motor nucleus of the primordial brain and forms the nucleus ruberin vertebrates.
From the first seven levels, essential components of the vertebrate brain developed. We call this initial structure the master brain of the segmented Bilateria.
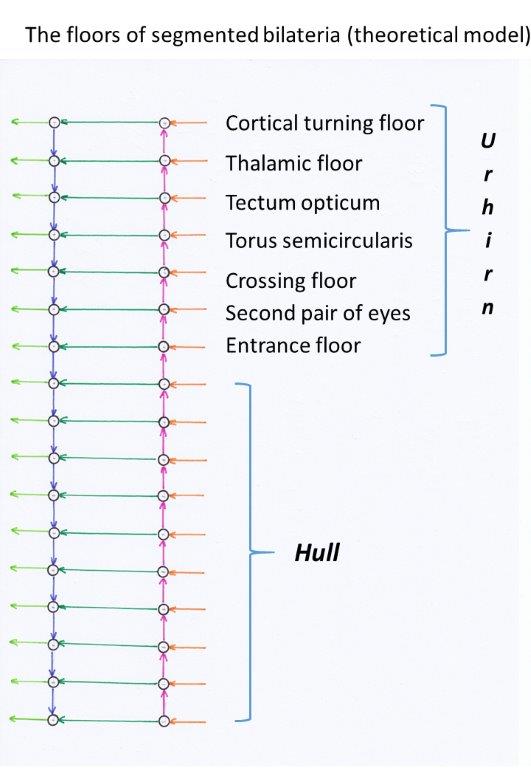
Figure 5 - Basic structures of the rope ladder nervous system
The bilaterally present mean centre of the seventh segment is formed by a nucleus called substantia nigra pars compactain vertebrates, which uses the transmitter dopamine.We assume that this mean centre was already present in the segmented bilateria whose line led to the vertebrates. Its earliest task is probably the regulation of the digestive and excretory system. Dopamine can still be detected in these substructures today.
In
addition to the input, output and mean value cores mentioned above and the
sensory and motor side-change cores, there were other cores, some of which will
be described in more detail at a later date, but some of which must remain
unmentioned, as this monograph would otherwise have become too extensive.
Reference should be made to a mean value nucleus that extended over several
segments and is referred to as the reticular format.
It runs like a net through the structures of the brain stem.
Theorem of the projections of the nucleus ruber
The bilaterally present nucleus ruber represents the nucleus of the primordial brain of the segmented Bilateria. It projects excitatory and descending to the motor neurons of the equilateral hemisphere via class 5 neurons. Furthermore, it projects via class 2 neuronsover a point-to-point connection via cross-commissures to the contralateral nucleus ruber into inhibitory interneurons there, whereby a contralateral inhibition of the projection neurons to the corresponding motoneurons of the opposite side is achieved.
The class 6 neurons in the nucleus ruber project into the mean centres of this floor and into the lower mean centres of the same side of the body.
Class 1 neurons in the nucleus ruber receive the excitatory output of the mean value centers of the different segments of the same side of the body.
The class 1 neuron signals arriving at the nucleus ruber caused a (relatively slight) pre-excitation of the downward projecting class 5 neurons and thus led to motor responses that were easier to induce.
On each side of the entrance level of the primordial brain there was one sensory and one motor side-change nucleus on each side of the body. We call the motoric side-changing nucleus nucleus olivaris.
Theorem of the nucleus olivaris as a side-changing nucleus of motor function
In the nucleus ruber there were two classes of projection neurons. Class 5 received the motor output of the primal brain and projected descending to the motor neurons. Class 2 also received the motor output of the primal brain from the neurons of class 5 and projected excitingly via the cross commissure to the motor side-changing nucleus, the nucleus olivaris. This receives the output of the ipsilateral nucleus ruber and initially projects excitatory to the opposite side into the nucleus ruber there, where it causes contralateral inhibition via inhibitory interneurons.
The bilaterally present nucleus olivaris was the motor side-changing nucleus of the entrance level of the primordial brain.
We recall that in the nucleus ruber there were also class 6 neurons that supplied the mean value systems with input. The mean value systems also had contralateral inhibition and were thus in neuronal competition with each other. For this purpose, the class 6 neurons also projected into an associated side-change nucleus. We assume that it fused with the nucleus olivaris, since both were very close neighbours and had the same tasks.
Thus, the nucleus olivaris contained a nucleus portion that projected the class 6 mean signals from the ipsilateral side to the contralateral side, where these also served the contralateral inhibition - this time of the class 6 mean signals. This mean value projection produced an output in the nucleus olivaris which was a significant input for the later Pontocerebellum.
The interneurons that caused the contralateral inhibition in the respective nucleus ruber separated in the course of evolution and formed their own neuronal nucleus, which we call the Purkinje nucleus.
Theorem of the origin of the Purkinje nucleus
In the nucleus ruber there were two subclasses of inhibitory interneurons. One class caused the lateral inhibition to enhance contrast, its input came from the ipsilateral side. The second class caused the contralateral inhibition of the projection neurons, their input came from the lateral change nucleus of the contralateral side. In the course of time, a spatial separation of these two neuron classes also occurred. The neurons for lateral inhibition remained in the nucleus as interneurons, while the neurons for contralateral inhibition segregated and formed their own nucleus over a longer evolutionary period, which we call the Purkinjekern in honor of the physiologist Johann Evangelista Purkinje. In the course of this cleavage, they changed from interneurons to projection neurons, as their axons now became significantly longer.
We therefore note a development in which, after a specialization of neurons of a nucleus on different tasks in the course of evolution, a splitting of such a nucleus occurs in such a way that the new sub-nuclei divide the functions of the division of labour among themselves. Each of the subnuclei specializes in a separate subtask. The subcores are also spatially separated in the course of evolution. We call such newly emerging subkernels descendants of the original nucleus.
The Purkinje nucleus is a descendant of the inhibitory interneurons of the nucleus ruber. It is a switching nucleus that switches the input from an excitatory to an inhibitory transmitter (glutamate->GABA ).
A small part of the Purkinje nuclei served to transmit the mean value signals of class 6, which, in addition to the class 5 signals, also arrived in the nucleus ruber and were included in the contralateral inhibition. In later evolutionary times it developed into the Pontocerebellum.
Theorem of the function of the Purkinje nucleus
The Purkinje core was used for motor contralateral inhibition at brainstem level. All sensory trunk signals of one side of the body that reached the brainstem via the entrance nucleus ascended to the individual levels of the primordial brain, were transmitted to the motor side on commissure class 3 neurons and reached the nucleus ruber in descending order. Each nucleus ruber projected on the one hand to the motor neurons of the trunk, also with the interposition of projection neurons of class 5 and via neurons of class 6 to the mean value centres. On the other hand, each nucleus ruber projected via class 2 neurons into the ipsilateral motor lateral change nucleus, the nucleus olivaris, which excitedly projected into the contralateral purkinje nucleus. The Purkinje core switched the signals to the inhibitory transmitter GABA and induced contralateral inhibition in the contralateral nucleus ruber via a 1:1 projection.
We decide to combine segments three to seven of the primordial brain into a structure that we call the brain stem.Then, contralateral inhibition at brainstem level wasrealized in the seventh segment by two nuclei, the nucleus olivaris and the purkinje nucleus. Independently of this, however, there was also the previous contralateral inhibition at the segment level.
Theorem of realization of contralateral motor inhibition at brainstem level
The neurons of the motor center of the seventh segment of each half of the body formed the nucleus ruber and projected excitatory to the ipsilateral nucleus olivaris. The nucleus olivaris in turn projected excitatory to the nucleus Purkinje, which switched these signals to the inhibitory transmitter GABA and projected inhibitory into the contralateral nucleus ruber, where in a point-to-point projection the contralateral inhibition of the corresponding contralateral signal took place.
The point-to-point projection essentially included the signals of the class 5 neurons, but a small part was reserved for the mean value signals of the class 6 neurons.
The Purkinje core of the early segmented Bilateria, whose line led to the vertebrates, was to develop into cerebellum barkin the course of evolution. From the signal projection of the class 5 neurons, the vestibulo- and spinocerebellum formed, the mean projection of the class 6 led to the formation of the pontocerebellum.
Before that, however, a reconstruction of the vestibular sense took place, which will be described later.
It seems that in the seventh segment a contralateral motor inhibition but no contralateral sensory inhibition developed, or that it was reduced in the course of evolution. In this segment, the incoming trunk signals were initially predominantly motor signals from the muscle spindles. For this type of signal, contralateral inhibition was already realized by the Purkinje core, so it was not necessary to use it on the sensory side.
However, this particularity did not apply to the head segments. There, contralateral inhibition developed, which was mainly based on the sensory signals of the head receptors, while motor contralateral inhibition apparently did not become significant.
An important sensory side-change nucleus of the visual tier is the nucleus isthmi (nucleus parabigeminalis of mammals), "which is reciprocally and topographically, i.e. with a point-to-point assignment, connected to the tectum (colliculi superiores of mammals) in all vertebrates and coordinates the activity of the two tectum hemispheres, for example in connection with the orientation of eye, head and hand movements and with depth perception"[48].
Quite inconspicuous - at least initially - are the connections of each sensory centre to the mean centre of the same segment of the same side of the body, because theoretically only one magnocellular mean neuron with its axon was needed to transmit a mean value, even if, for reasons of transmission security, several mean neurons formed a small axon bundle with reserve axons. In the course of evolution, both specializations of the mean nuclei- recognizable by the use of different transmitters - and the reduction of mean nuclei in the trunk segments of the body occurred.
The neural system consisting of two sensory and two motor strands is also called the four-strand system or tetraneural nervous system.Annelids are an example of segmented bilateria, which originally had such a nervous system, although it has been simplified by reduction.
Several groups of the Bilateria formed paired structures on their segments, which could be used for locomotion. The annelids developed paired lateral stubby feet (paradopia) on the segments. The arthropods (Euarthropoda) produced a pair of extremities on each of their trunk segments.
In the course of evolution, some of the segmented bilateria described above have been simplified. If, for example, locomotion (in mud and sand) was mainly achieved by alternating contraction of the ring muscles on the one hand and the longitudinal muscles on the other hand, contralateral inhibition hardly played a role. Then the two parallel rope ladder systems of the two halves of the body could merge.This possibly meant the complete merging of the left and right sensory center and also the left and right motor center in each segment without abandoning the separation of functionality. This is because the left sensory centre still received the sensory signals from the left half of the body and transmitted them to the neurons of the motor centre of the left half, from where they controlled the motor neurons of the left half of the body. Likewise, the signaling pathways of the right half of the body remained self-sufficient.
It is also possible to unite the left and right rope ladder systems into one, without abandoning the functional separation, because the contralateral inhibition became an important part of locomotion. In this case the leg pairs of a segment were not moved synchronously to each other but asynchronously, at best even in opposite directions. This was particularly effective when there were only a few segments and the total number of legs was only four, six or eight. Then an asynchronous control of the legs of the same segment was inevitably counter-rotating. The required contralateral inhibition of the contralateral leg led to the fanning out of a possibly folded rope ladder system, so that the nervous system became tetraneural again at these segment heights.
In the beginning the number of muscles was very low anyway. Only when the number of muscles increased strongly - for example in fish - was the spatial separation of the left and right rope ladder system particularly necessary, since here the number of cross connections for the required contralateral inhibition also increased just as strongly.
Of course, the question arises as to what evolutionary advantage the bilateral segment animals had. The greatest advantage was the possibility of effective locomotion in water, triggered by stimulation of the receptors. One reason was the frequency multiplication of the action potentials of receptors due to the ladder structure of the system, the second was the contralateral inhibition, and the third was the noticeable time delay of action potentials on non-markless axons.
Theorem of frequency multiplication
If a receptor in a segment of a tetraneural nervous system was stimulated to generate an action potential which was to act on a muscle located in this segment, the ladder structure in each segment located above it triggered a further action potential. The ascending action potential, passed on by the connective neurons, generated an action potential in each segment level, which was passed on via the commissures and which was supplied in the motor strand to the muscle in the original segment. This corresponds to a frequency multiplication. The closer a segment was located to the end of the body, the greater was the frequency multiplication of the sensory signals that reached the muscles in this segment.
Thus, weak and short-term effects on receptors in the segment animals led to significantly stronger signal responses. The effect was stronger with tail-side stimulation than with head-side stimulation. The stronger signal responses were already sufficient to generate muscle contractions. Therefore, segment animals were superior to the others with regard to their reaction speed.
Theorem of more effective movement through contralateral inhibition
The contralateral escapement allowed for a wiggly movement that led to a higher
speed. This became particularly effective when fin-like elements used the water
resistance.
During locomotion with limbs, their partial elements were included in the
contralateral inhibition, so that the motor opponent was prevented by inhibition
from preventing the movement by its own contraction.
Theorem of the importance of time delay on non-markless fibers for motion
A continuous wave of contractions and decontractions along the body was essential for a meandering movement and was made possible by the time-delayed transfer of excitation from one segment to the next. The required time delay was caused by the low velocity of propagation of action potentials along non-markless axons. The time delay was also partly responsible for the contralateral inhibition of motor opponents.
We know that the axons were non-markless at that time, myelin developed much later. So the velocity of propagation along the axons was relatively slow. The excitation of the neurons in one segment was passed on with a time delay, on the one hand excitatory to the segment above, on the other hand via the cross-commissure and inhibitory interneurons to the neurons of the opposite side in the same segment. This resulted in a snake-like movement of the body, which was excellently suited for forward movement in water. The contralateral inhibition allowed the contraction of a muscle because the motor counter muscle remained relaxed by the inhibition and gave way to the contraction initiated on the opposite side.
The first segmented bilaterias had a hydroskeleton. The exoskeleton, which did not grow with the moulting animals and was stripped off and newly formed by moulting at certain intervals, for example in insects or crustaceans, achieved a higher strength. During the moulting phase, additional segments were often formed.
An inner skeleton could provide greater strength combined with good mobility. Here the chorda dorsalis could prove its worth. All vertebrates are chordates, i.e. they possess a mobile support rod (chorda dorsalis), either during embryonic development or afterwards, from which the vertebrate spine of the vertebrates emerged. In aquatic chordates, much greater forces could be absorbed to enable locomotion in water.
In connection with the development of an internal skeleton, it was necessary to improve the protection of the body surface on the one hand through the rapid analysis of external influences by allowing the recognition of a sometimes extremely high number of receptors. On the other hand, a fast motor reaction was necessary to prevent an attack by predators, for example by escape or counterattack. In this case, a stable internal skeleton was ideally suited to convert strong tensile stresses of muscles into movements.
Theorem of the transition from hydroskeleton to internal skeleton in choral data
An internal skeleton made it possible to use much stronger muscle tensions to generate movement. While an exoskeleton covered large parts of the body surface, the inner skeleton covered the body surface uncovered, so that a high number of receptors had to be developed to monitor the body surface. This led to a significant increase in the number of neurons and the axons connecting them in the nervous system. The omission of an exoskeleton also simplified the increase in size of living organisms, which now also led to the formation of new muscle systems. This also increased the number of motor neurons and the output neurons of the motor centres of the different segments.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan