Gehirntheorie der Wirbeltiere
für das Human Brain Projekt der Europäischen Union
ISBN 978-3-00-064888-5
3.4. Die Grundstrukturen im Urhirn der segmentierten Bilateria
Die oberste Kommissur, die im Kopfsegment das sensorische Zentrum mit dem motorischen verbindet, bezeichnen wir künftig als cortikale Wendeetage. Das sensorische und motorische Zentrum des Kopfsegmentes betrachten wir als Teil der cortikalen Wendeetage. Aus ihr ging in einem längeren Entwicklungsprozess der Cortex der Wirbeltierehervor, der sich wiederum in einen motorischen und einen sensorischen Cortex unterteilen lässt, wobei der sensorische Cortex nach Modalitäten wohlgeordnet ist. Dies wird in den folgenden Kapiteln nachgewiesen. Der Mittelwertkern der olfaktorischen cortikalen Etage ist das Septum.
Die analoge Struktur, die bereits aus der Verschmelzung der kopfseitigen Segmente entstand, bezeichnen wir als thalamische Wendeetage. Aus ihr entwickelte sich das thalamische System der Wirbeltiere. Das ebenfalls bilateral vorhandene Mittelwertzentrum dieser Etage wird von einem Mittelwertkern gebildet, der bei Wirbeltieren als Nucleus subthalamicusbezeichnet wird. Diesen Namen werden wir bereits hier verwenden.
Das sensorische und das motorische Zentrum der dritten Etagebildet zusammen mit den zugehörigen Kommissuren eine Struktur, die wir als optisches Tectumbezeichnen. Es stellt (von oben gezählt) die dritte Wendeetagedar. In ihr kommt es zu Signalwechselwirkungen zwischen motorischen und visuellen Signalen.
Der vierten Wendeetageordnen wir eine Struktur zu, die als Torus semicircularis bezeichnet wird. Hier wirken die Signale des Vestibularsinnes und (falls vorhanden) die des Seitenliniensystems auf die motorischen Signale ein.
Die fünfte Wendeetagereservieren wir für eine Struktur, die später beschrieben werden wird und die wir als Kreuzungsetagebezeichnen werden.
Die sechste Wendeetagereservieren wir für ein weiteres Augenpaar. Hierbei postulieren wir, dass hier keine Linsenaugen, sondern konvexe Facettenaugenverwendet werden. In diese Etage ordnen wir auch das optische Mittelwertsystem ein, welches den Nucleus suprachiasmaticusmit Signalen versorgt.
Die siebente Wendeetage(von oben) stellt das Eingangssystemoder die Eingangsetagedar. Sein sensorisches Zentrum stellt den Eingangskerndes Urhirns dar und empfängt die Rezeptorsignale der Rumpfsegmente. Dieser Eingangskern spaltete sich bei den Wirbeltieren in zwei Unterkerne auf, die als Nucleus cuneatusund Nucleus gracilisbezeichnet werden. Der Erstere erhält die Rezeptorsignale der oberen Rumpfhälfte, der andere die der unteren. Gleichzeitig ist dieses Segment auf der motorischen Seite die Ausgangsetage.
Das motorische Zentrum des siebenten Segmentes stellt den motorischen Ausgangskerndes Urhirns dar und bildet bei den Wirbeltieren den Nucleus ruber.
Aus den ersten sieben Etagen entwickelten sich wesentliche Komponenten des Gehirns der Wirbeltiere. Wir bezeichnen diese anfängliche Struktur als Urhirn der segmentierten Bilateria.
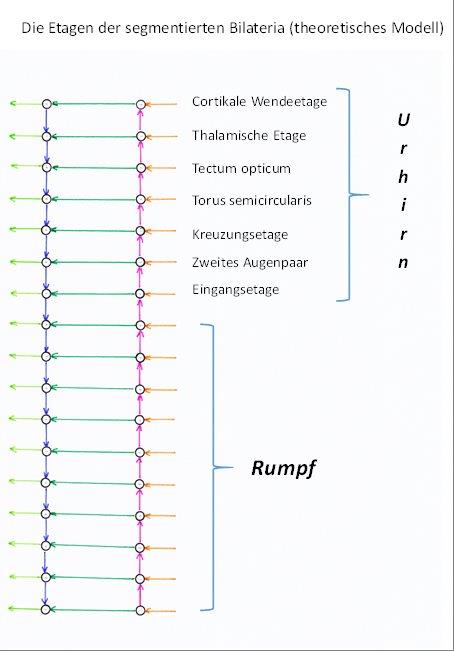
Abbildung5- Grundstrukturen des Strickleiternervensystem
Das bilateral vorhandene Mittelwertzentrum des siebenten Segmenteswird von einem Kern gebildet, der bei Wirbeltieren als Substantia nigra pars compactabezeichnet wird und den Transmitter Dopaminverwendet. Wir unterstellen, dass dieses Mittelwertzentrum bereits bei den segmentierten Bilateria vorhanden war, deren Linie zu den Wirbeltieren führte. Seine frühste Aufgabe dürfte die Regulierung des Verdauungs- und Ausscheidungssystems sein. In diesen Substrukturen lässt sich noch heute Dopamin nachweisen.
Neben den genannten Eingangs-, Ausgangs- und Mittelwertkernen und den sensorischen und motorischen Seitenwechselkernen existierten weitere Kerne, die teils zu einem späteren Zeitpunkt genauer beschrieben werden, teils jedoch auch unerwähnt bleiben müssen, da diese Monografie sonst zu umfangreich geworden wäre. Hingewiesen werden sollte auf einen Mittelwertkern, der sich über mehrere Segmente erstreckte und als Formatio reticularisbezeichnet wird. Er durchzieht netzartig die Strukturen des Hirnstamms.
Theorem der Projektionen des Nucleus ruber
Der bilateral vorhandene Nucleus ruberstellt den Ausgangskern des Urhirns der segmentierten Bilateria dar. Er projiziert über Neuronen der Klasse 5 erregend und absteigend zu den Motoneuronen der gleichseitigen Körperhälfte. Weiterhin projiziert er mittels der Neuronen der Klasse 2über eine Punkt-zu-Punkt-Verbindung über Kreuzkommissuren zum kontralateralen Nucleus ruberin dortige hemmende Interneuronen, wodurch eine kontralaterale Hemmungder Projektionsneuronen zu den korrespondierenden Motoneuronen der Gegenseite erreicht wird.
Die Neuronen der Klasse 6 im Nucleus ruber projizieren in die Mittelwertzentren dieser Etage und in die tiefer gelegenen Mittelwertzentren der gleichen Körperseite.
Neuronen der Klasse 1 im Nucleus ruber empfangen den erregenden Output der Mittelwertzentren der verschiedenen Segmente der gleichen Körperseite.
Die im Nucleus ruber eintreffenden Signale der Neuronenklasse 1 bewirkten eine (relativ geringfügige) Vorerregung der abwärts projizierenden Neuronen der Klasse 5 und führten so zu leichter zu bewirkenden motorischen Reaktionen.
Auf jeder Seite der Eingangsetage des Urhirns gab es je einen sensorischen und einen motorischen Seitenwechselkern auf jeder Körperseite. Der motorische Seitenwechselkern wird von uns als Nucleus olivaris bezeichnet.
Theorem des Nucleus olivaris als Seitenwechselkern der Motorik
Im Nucleus ruber gab es zwei Klassen von Projektionsneuronen. Die Klasse 5 empfing den motorischen Output des Urhirns und projizierte absteigend zu den Motoneuronen. Die Klasse 2 empfing von den Neuronen der Klasse 5 ebenfalls den motorischen Output des Urhirns und projizierte erregend über die Kreuzkommissur zum motorischen Seitenwechselkern, dem Nucleus olivaris. Dieser empfängt den Output des ipsilateralen Nucleus ruber und projiziert anfänglich erregend zur Gegenseite in den dortigen Nucleus ruber, wo er über hemmende Interneuronen die kontralaterale Hemmung bewirkt.
Der bilateral vorhandene Nucleus olivaris war der motorische Seitenwechselkern der Eingangsetage des Urhirns.
Wir erinnern uns daran, dass es im Nucleus ruber auch die Neuronen der Klasse 6 gab, die die Mittelwertsysteme mit Input versorgten. Auch die Mittelwertsysteme besaßen eine kontralaterale Hemmung und standen so in neuronaler Konkurrenz zueinander. Dazu projizierten die Neuronen der Klasse 6 auch in einen zugehörigen Seitenwechselkern. Wir gehen davon aus, dass er mit dem Nucleus olivaris verschmolz, da beide sehr eng benachbart waren und gleiche Aufgaben hatten.
Der Nucleus olivaris enthielt also einen Kernanteil, der die Mittelwertsignale der Klasse 6 der ipsilateralen Seite zur kontralateralen Seite projizierte, wo diese ebenfalls der kontralateralen Hemmung - dieses Mal der Mittelwertsignale der Klasse 6 - dienten. Diese Mittelwertprojektion brachte im Nucleus olivaris einen Output hervor, der einen wesentlichen Input für das spätere Pontocerebellumdarstellte.
Die Interneuronen, die im jeweils Nucleus ruber die kontralaterale Hemmung bewirkten, separierten sich im Laufe der Evolution und bildeten einen eigenen neuronalen Kern, den wir als Purkinjekern bezeichnen.
Theorem der Entstehung des Purkinjekerns
Im Nucleus ruber gab es zwei Unterklassen von hemmenden Interneuronen. Eine Klasse bewirkte die laterale Hemmung zur Kontrastverstärkung, ihr Input entstammte der ipsilateralen Seite. Die zweite Klasse bewirkte die kontralaterale Hemmung der Projektionsneuronen, ihr Input entstammte dem Seitenwechselkern der kontralateralen Seite. Hier trat im Verlaufe der Zeit auch eine räumliche Trennung dieser zwei Neuronenklassen auf. Die Neuronen für die laterale Hemmung verblieben im Kern als Interneuronen, während sich die Neuronen für die kontralaterale Hemmung absonderten und in einem längeren evolutionären Zeitraum einen eigenen Kern bildeten, den wir zu Ehren des Physiologen Johann Evangelista Purkinjeals Purkinjekernbezeichnen. Im Zuge dieser Abspaltung wandelten sie sich von Interneuronen zu Projektionsneuronen, da ihre Axone nun deutlich länger wurden.
Wir nehmen daher eine Entwicklung zur Kenntnis, bei der nach einer Spezialisierung von Neuronen eines Kerns auf unterschiedliche Aufgaben im Verlaufe der Evolution eine Aufspaltung eines solchen Kerns derart erfolgt, dass die neuen Teilkerne die arbeitsteiligen Funktionen unter sich aufteilen. Jeder der Subkerne spezialisiert sich auf eine separate Teilaufgabe. Die Subkerne werden im Verlaufe der Evolution auch räumlich getrennt. Wir bezeichnen solche neu entstehenden Subkerne als Abkömmlinge des ursprünglichen Kerns.
Der Purkinjekern ist ein Abkömmling der hemmenden Interneuronen des Nucleus ruber. Er ist ein Umschaltkern, der den Input von einem erregenden auf einen hemmenden Transmitter umschaltet (Glutamat->GABA).
Ein kleiner Teil der Purkinjekerns diente der Weiterleitung der Mittelwertsignale der Klasse 6, die neben den Signalen der Klasse 5 ebenfalls im Nucleus ruber eintrafen und in die kontralaterale Hemmung einbezogen wurden. In späterer Evolutionszeit entwickelte er sich zum Pontocerebellum.
Theorem der Funktion des Purkinjekerns
Der Purkinjekern diente der motorischen kontralateralen Hemmung auf Hirnstammebene. Alle sensorischen Rumpfsignale einer Körperseite, die über den Eingangskern den Hirnstamm erreichten, stiegen in die einzelnen Etagen des Urhirns auf, wurden auf Kommissurneuronen der Klasse 3 zur motorischen Seite weitergeleitet und erreichten absteigend den Nucleus ruber. Jeder Nucleus ruber projizierte einerseits zu den Motoneuronen des Rumpfes, auch unter Zwischenschaltung von Projektionsneuronen der Klasse 5 sowie über Neuronen der Klasse 6 zu den Mittelwertzentren. Andererseits projizierte er über Neuronen der Klasse 2 in den ipsilateralen motorischen Seitenwechselkern, den Nucleus olivaris, der erregend in den kontralateralen Purkinjekern projizierte. Der Purkinjekern schaltete die Signale auf den hemmenden Transmitter GABA um und bewirkte im kontralateralen Nucleus ruber über eine 1:1-Projektion die kontralaterale Hemmung.
Wir entschließen uns, die Segmente drei bis sieben des Urhirns zu einer Struktur zusammenzufassen, die wir als Hirnstammbezeichnen. Dann wurde die kontralaterale Hemmung auf Hirnstammebeneim siebenten Segment durch zwei Kerne realisiert, den Nucleus olivaris und den Purkinjekern. Unabhängig davon gab es jedoch auch die bisherige kontralaterale Hemmung auf Segmentebene.
Theorem der Realisierung der kontralateralen motorischen Hemmung auf Hirnstammebene
Die Neuronen des motorischen Zentrums des siebenten Segments einer jeden Körperhälfte bildeten den Nucleus ruber und projizierten erregend zum ipsilateralen Nucleus olivaris. Der Nucleus olivaris seinerseits projizierte erregend zum Nucleus Purkinje, der diese Signale auf den hemmenden Transmitter GABA umschaltete und hemmend in den kontralateralen Nucleus ruber projizierte, wo in einer Punkt-zu-Punkt-Projektiondie kontralaterale Hemmung des zugehörigen kontralateralen Signals erfolgte.
Die Punkt-zu-Punkt-Projektion umfasste im Wesentlichen die Signale der Neuronen der Klasse 5, jedoch war ein geringer Teil den Mittelwertsignalen der Neuronen der Klasse 6 vorbehalten.
Der Purkinjekernder frühen segmentierten Bilateria, deren Linie zu den Wirbeltieren führte, sollte sich im Verlauf der Evolution zur Cerebellumrindeentwickeln. Aus der Signalprojektion der Neuronen der Klasse 5 bildete sich das Vestibulo- und das Spinocerebellum, die Mittelwertprojektion der Klasse 6 führte zur Entstehung des Pontocerebellums.
Zuvor jedoch fand ein Umbau des Vestibularsinnes statt, der noch zu beschreiben sein wird.
Es scheint, dass sich im siebenten Segment zwar eine kontralaterale motorische, jedoch keine kontralaterale sensorische Hemmung herausgebildet hat, oder dass sie im Verlaufe der Evolution zurückgebildet wurde. In diesem Segment waren die eintreffenden Rumpfsignale anfänglich vorwiegend motorische Signale der Muskelspindeln. Für diese Signalart wurde die kontralaterale Hemmung bereits vom Purkinjekern realisiert, daher konnte auf der sensorischen Seite darauf verzichtet werden.
In den Kopfsegmenten traf diese Besonderheit jedoch nicht zu. Dort bildete sich eine kontralaterale Hemmung aus, die vorwiegend auf den sensorischen Signalen der Kopfrezeptoren beruhte, während eine motorische kontralaterale Hemmung offenbar keine Bedeutung erlangte.
Ein wichtiger sensorischer Seitenwechselkern der visuellen Etage ist der Nucleus isthmi (Nucleus parabigeminalis der Säuger), -der bei allen Wirbeltieren reziprok und topografisch , das heißt mit einer Punkt-zu-Punkt-Zuordnung, mit dem Tectum (Colliculi superiores der Säuger) verbunden ist und die Aktivität der beiden Tectum-Hemisphären koordiniert, etwa im Zusammenhang mit der Orientierung von Augen-, Kopf- und Handbewegungen und mit der Tiefenwahrnehmung.-[48]
Recht unauffällig - jedenfalls anfänglich - sind die Verbindungen jedes sensorischen Zentrums zum Mittelwertzentrum des gleichen Segments der gleichen Körperseite, denn zur Übertragung eines Mittelwertes war theoretisch nur ein magnozellulares Mittelwertneuronmit seinem Axon nötig, auch wenn aus Gründen der Übertragungssicherheit mehrere Mittelwertneuronen ein kleines Axonbündel mit Reserveaxonen bildeten. Im Verlaufe der Evolution kam es sowohl zu Spezialisierungen der Mittelwertkerne- erkennbar an der Verwendung unterschiedlicher Transmitter - als auch zur Reduzierung der Mittelwertkerne in den Rumpfsegmenten des Körpers.
Das aus zwei sensorischen und zwei motorischen Strängen bestehende neuronale System wird auch als Vierstrangsystemoder tetraneurales Nervensystembezeichnet. Ringelwürmer(Anneliden) sind ein Beispiel für segmentierte Bilateria, die ursprünglich ein solches Nervensystem besaßen, auch wenn es teils durch Reduktion vereinfacht worden ist.
Etliche Gruppen der Bilateria bildeten an ihren Segmenten paarige Strukturen, die der Fortbewegung dienen konnten. Die Ringelwürmer oder auch Gliederwürmer (Anneliden) entwickelten an den Segmenten paarige laterale Stummelfüße (Paradopien). Die Gliederfüßer (Euarthropoda) brachten an ihren Rumpfsegmenten je ein Paar Extremitäten hervor.
Im Verlaufe der Evolution konnte es bei einem Teil der beschriebenen segmentierten Bilateria zu gewissen Vereinfachungen kommen. Erfolgte beispielsweise die Fortbewegung (im Schlamm und Sand) vorwiegend durch wechselweise Kontraktion einerseits der Ringmuskulatur und andererseits der Längsmuskulatur, so spielte eine kontralaterale Hemmung kaum eine Rolle. Dann konnte es zur Verschmelzungder zwei parallelen Strickleitersysteme der beiden Körperhälften kommen. Dies bedeutete möglicherweise die komplette Verschmelzung des linken und des rechten sensorischen Zentrums und ebenso des linken und des rechten motorischen Zentrums in jedem Segment, ohne dass die Trennung der Funktionalität aufgegeben wurde. Denn das linke sensorische Zentrum empfing nach wie vor die sensorischen Signale der linken Körperhälfte und vermittelte sie an die Neuronen des motorischen Zentrums der linken Hälfte, von wo aus sie die Motoneuronen der linken Körperhälfte ansteuerten. Ebenso blieben die Signalwege der rechten Körperhälfte autark erhalten.
Ebenso möglich ist die räumliche Vereinigung des linken und des rechten Strickleitersystems zu einem gemeinsamen, ohne dass auch hier die funktionale Trennung aufgegeben wurde, weil die kontralaterale Hemmung wichtiger Bestandteil der Fortbewegung wurde. Hier wurden dann die Beinpaare eines Segmentes nicht synchron zueinander bewegt, sondern asynchron, bestenfalls sogar gegenläufig. Dies war besonders effektiv, wenn nur wenige Segmente vorhanden waren und die Gesamtzahl der Beine nur vier, sechs oder acht betrug. Dann war eine asynchrone Ansteuerung der Beine des gleichen Segments zwangsläufig gegenläufig. Die erforderliche kontralaterale Hemmung des kontralateralen Beins führte zum Auffächern eines möglicherweise zusammengelegten Strickleitersystems, so dass das Nervensystem auf diesen Segmenthöhen wieder tetraneural wurde.
Anfänglich war die Anzahl der Muskeln sowieso sehr niedrig. Erst als die Muskelanzahl stark anstieg - so etwa bei den Fischen - war die räumliche Trennung des linken und des rechten Strickleitersystems besonders notwendig, da hier auch die Anzahl der Kreuzverbindungen zur erforderlichen kontralateralen Hemmung ebenso stark anstieg.
Natürlich ergibt sich die Frage, welchen evolutionären Vorteil die bilateralen Segmenttiere besaßen. Der größte Vorteil lag in der Möglichkeit der effektiven Fortbewegung im Wasser, ausgelöst durch Reizeinwirkung auf die Rezeptoren. Eine Ursache war die Frequenzvervielfachungder Aktionspotentiale von Rezeptoren durch die Leiterstruktur des Systems, die zweite die kontralaterale Hemmung, die dritte war die merkliche Zeitverzögerungder Aktionspotentiale auf marklosen Axonen.
Theorem der Frequenzvervielfachung
Wurde in einem Segment eines tetraneuralen Nervensystems ein Rezeptor zur Generierung eines Aktionspotentials angeregt, welches auf einen in diesem Segment befindlichen Muskel einwirken sollte, kam es durch die Leiterstruktur in jedem darüber befindlichen Segment zur Auslösung eines weiteren Aktionspotentials. Das aufsteigende Aktionspotential, weitergebenen durch die Konnektivneuronen, generierte in jeder Segmentebene ein über die Kommissuren weitergegebenes Aktionspotential, welches im motorischen Strang zum Muskel im ursprünglichen Segment zugeführt wurde. Dies entspricht einer Frequenzvervielfachung. Je näher ein Segment zum Körperende gelegen war, umso größer war die Frequenzvervielfachung der sensorischen Signale, die von diesem Segment aus die in ihm befindlichen Muskeln erreichten.
So kam es, dass bei den Segmenttierenschwache und kurzzeitige Einwirkungen auf Rezeptoren zu deutlich stärkeren Signalantworten führten. Die Wirkung war bei schwanzseitiger Reizung stärker als bei kopfseitiger. Die stärkeren Signalantworten reichten bereits zur Generierung von Muskelkontraktionen aus. Daher waren Segmenttiere hinsichtlich ihrer Reaktionsgeschwindigkeitden anderen überlegen.
Theorem der effektiveren Bewegungdurch kontralaterale Hemmung
Die kontralaterale Hemmung erlaubte eine schlängelnde Bewegung, die zu einer höheren Geschwindigkeit führte. Besonders effektiv wurde diese, wenn flossenartige Elemente den Wasserwiderstand nutzten.
Bei der Fortbewegung mit Gliedmaßen wurden deren Teilelemente in die kontralaterale Hemmung einbezogen, so dass der motorische Gegenspieler durch Hemmung daran gehindert wurde, die Bewegung durch eigene Kontraktion zu verhindern.
Theorem der Bedeutung der Zeitverzögerung auf marklosen Fasern für Bewegungen
Eine entlang des Körpers fortlaufende Welle von Kontraktionen und Dekontraktionen war für eine schlängelnde Bewegung unabdingbar und wurde ermöglicht durch die zeitversetzte Weitergabe der Erregungen von einem Segment zum benachbarten. Die erforderliche Zeitverzögerung wurde durch die geringe Ausbreitungsgeschwindigkeit von Aktionspotentialen entlang markloser Axone verursacht. Ebenso mitverantwortlich war die Zeitverzögerung bei der kontralateralen Hemmung der motorischen Gegenspieler.
Wir wissen, dass die Axone zur damaligen Zeit marklos waren, Myelin entwickelte sich viel später. Daher war die Ausbreitungsgeschwindigkeit entlang der Axone relativ gering. Die Erregung der Neuronen in einem Segment wurde zeitverzögert weitergegeben, einerseits erregend an das darüber liegende Segment, andererseits über die Kreuzkommissur und hemmende Interneuronen an die Neuronen der Gegenseite im gleichen Segment. Dadurch kam eine schlangenartige Bewegung des Körpers zustande, die im Wasser vorzüglich geeignet war für eine Vorwärtsbewegung. Die kontralaterale Hemmung erlaubte die Kontraktion eines Muskels, weil der motorische Gegenspielermuskel durch die Hemmung entspannt blieb und der eingeleiteten Kontraktion auf der Gegenseite nachgab.
Die ersten segmentierten Bilateria hatten ein Hydroskelett. Eine höhere Festigkeit erreichte das Exoskelett, welches bei den Häutungstieren nicht mitwuchs und in gewissen Abständen durch Häutung abgestreift und neu gebildet wurde, etwa bei Insekten oder Krebsen. Während der Häutungsphase wurden oft auch weitere Segmente gebildet.
Eine höhere Festigkeit, gepaart mit einer guten Beweglichkeit, konnte ein Innenskelettbieten. Hier konnte sich die Chorda dorsalis bewähren. Alle Wirbeltiere sind Chordaten, besitzen also entweder während der Embryonalentwicklung oder auch danach einen beweglichen Stützstab (Chorda dorsalis), aus dem die Wirbelsäule der Wirbeltiere hervorging. Bei den wasserbewohnenden Chordaten konnten so viel größere Kräfte aufgenommen werden, um eine Fortbewegung im Wasser zu ermöglichen.
Verbunden mit der Herausbildung eines Innenskeletts war die Notwendigkeit, den Schutz der Körperoberfläche einerseits durch die schnelle Analyse von äußeren Einwirkungen zu verbessern, indem eine teils extrem hohe Anzahl von Rezeptoren solche zu erkennen gestattete. Andererseits war eine schnelle motorische Reaktion nötig, um einem Angriff von Fressfeinden etwa durch Flucht oder Gegenangriff zuvorzukommen. Hier war ein stabiles Innenskelett bestens geeignet, starke Zugspannungen von Muskeln in Bewegungen umzusetzen.
Theorem des Übergangs vom Hydroskelett zum Innenskelett bei den Chordaten
Ein Innenskelett ermöglichte es, viel stärkere Muskelspannungen zur Erzeugung von Bewegungen einzusetzen. Während ein Exoskelett große Teile der Körperoberfläche bedeckte, war beim Innenskelett die Körperoberfläche unbedeckt, so dass eine hohe Anzahl von Rezeptoren zur Überwachung der Körperoberfläche entwickelt werden musste. Dadurch stieg die Anzahl der Neuronen und der sie verbindenden Axone im Nervensystem deutlich an. Der Wegfall eines Exoskeletts vereinfachte auch die Größenzunahmen der Lebewesen, die sich nun ebenfalls in der Herausbildung von neuen Muskelsystemen äußerte. Damit stieg ebenso die Anzahl der Motoneuronen und der Outputneuronen der motorischen Zentren der verschiedenen Segmente.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan