Vertebrate brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-064888-5
3.6 The topological well-being in the head area
The visual topology of compound eyes
Segmented bilateria often have sensory organs that are distributed over many segments. More highly organised ones usually have both lens eyes (median eyes) andcompound eyes (compound eyes) on their head segments, sometimes the compound eyes are also resolved into single eyes. We assume here that compound eyes represent the original eye variant. Therefore, we investigate the question why lens eyes developed later. But first we analyze the imaging properties of both eye types.
Theorem of correct and upright imaging in convex compound eyes
In convex compound eyes,an upright and laterally correct image is formed on the visual receptor surface, the resolution of which increases with the number of facets.
Here we assume that the surface of the compound eyes is convex, i.e. curved outwards.
Theorem of inverted and upside-down imaging with concave compound eyes
In concave compound eyes, a laterally inverted and upside down image is formed on the retinal surface, the resolution of which increases with the number of receptors.
A facet eye is defined as concave if its facets form a concave surface that is curved inwards. If the depression of the surface is large, we can speak of a pit eye. It is not difficult to imagine the transition from a pit eye to a lens eye.
Theorem of upside-down and inverted imaging in lens eyes
In lens eyes, a laterally inverted and upside-down image is formed on the retinal surface, the resolution of which increases with the number of receptors.
The actual benefit of visual images is based on the property of light-sensitive receptors to transform light effects into sequences of action potentials. Since the earliest living creatures lived in water, which (without foreign substances and at shallower depths) was itself very translucent, the receptors reacted best to dark objects. The less light there was, the higher the rate of fire, and with increasing brightness it decreased. Therefore, the eyes initially detected dark objects.
We assume that these dark objects were initially interpreted as food. Had it been the other way round, the early animal life forms would have (possibly) died out because they fled from any food, as if every dark object was a predator.
How could the visual signals now trigger a movement towards the feed?
We must remember that the muscle tension receptors that ultimately act on the motor neurons projected in the neural tube in each segment onto a body image that was located on the outer surface of a half cylinder. In this body image, the segments of the body formed segment strips. One quarter cylinder each represented one half of the body of the bilateral animal.
The retina, on the other hand, was a circular area. This formed a circular body model of the visual receptors. In order for these to become motorically effective, this circular body model had to be converted into a semi-cylindrical one, which could be attached to the sensory body model of the muscle tension receptors on the outside. The site of this attachment was the tectum opticum. This attachment was possible if the circular retina was first transferred into a semicircle and then imaged onto a semicylinder. We will describe these two steps in more detail.
First, we define the visual mapping function, which should be a visual circular function because it is defined on a circle that has the size of the visual receptor area (retina). The function value should correspond to the current rate of fire in the area in question. This function is not continuous, but discrete. A function value is only present where a receptor is actually located.
Definition: visual circular function
The mapping of visual receptors to a circular area while maintaining their neighbourhood relationships defines a visual circular function. The function value results from the fire rate of the receptors. The function is only defined and discrete in the pixels of the receptors.
Considering the fact that in segmented bilaterias with separation of sensory and motor axons, the sensory input can only use half of a neural tube, we transform the visual circle function into a visual semicircle function by mapping the circle to a semicircle.
Definition: visual semi-circular function
The mapping of a visual circle function to a semicircle is called a visual semicircle function.
We now transfer the visual semicircle function into a visual mantle function defined on the mantle surface of a half cylinder. Each receptor located in the semicircular surface may send its axon to the assigned point of the mantle surface and excite a receiving neuron there at the same rate of fire as it has itself. The outer edge of the semicircle may be mapped on the top of the mantle surface, the centre of the circle on the bottom.
In the above-mentioned transfers from one form of representation to another, the topology should always be preserved, especially neighbourhood relations between pixels.
Definition: visual sheath function
We call the mapping of a visual semicircular function onto the outer surface of a half cylinder a visual mantle function.
Now we remember the theorem of segmented body images in the trunk area. In each segment there existed for each modality a body image on the mantle surface of a half cylinder. In the head area there is an analogous situation, because all existing axons also move into the second, the visual segment via connective connections.
There, overlapping occurs in that the mantle surface of the visual receptor neurons is attached to the outside of the previous mantle surfaces. Thus, the sensory axons of the trunk run inside and contact the corresponding mantle neurons, while the visual axons run outside and contact the visual mantle neurons.
Via interneurons, the incoming visual signals may now excite the sensory mantle neurons and imprint the visual excitation on them.
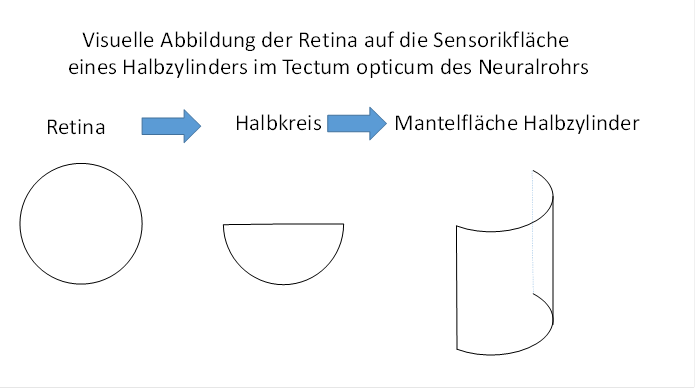
Figure 8 - Visual imaging of the retina in the optic tectum
What happens now if it is a compound eye?
Theorem of visual muscle control in compound eyes
In combination with an independent forward movement, a visual object located at the top of a compound eye leads to a contraction of the upper trunk muscles, so that the movement is in the direction of the object. An object at the bottom generates a contraction of the lower trunk muscles. A left object causes the left trunk muscles to contract, and a right object causes the right trunk muscles to contract. An object in the center of vision activates the tail muscles of the last segment, while an object at the edge of the field of view activates the muscles of the head side segments. A normal horizontal position, such as that of a fish, is required.
The centre of the field of view is assigned to the tail segment, the periphery of the field of view to the head segment. With increasing distance from the centre, the segment number of the segment increases, which receives the corresponding visual signals as control signals for its motoneurons.
We refer to such an image as a standard visual motor projection without lateral body reflection. This leads to an approach to visual objects so that they can ultimately be grasped with the mouth. The approaching movement is meandering by reciprocal oscillation caused by the contralateral inhibition and the time delay on the myelin-free class 2 cross commissures, and the corrective movements generated on the retina by the object image on the retina, which is also oscillating.
With this we can formulate the following theorem.
Theorem of standard visual motor projection in compound eyes with convex curvature
Facet eyes with convex curvature have a standard visual motor projection without lateral body reflection.
With lens eyes, the visual image would be reversed and vice versa under the same conditions, so the movements would be reversed. With this imaging topology, lens eyes would lead to active evasion of the food. Therefore, lenticular eyes cannot be used for foraging at this stage of evolution. Their training requires the development of an inverse imaging topology. This occurred much later and required a cause which will be described in the following sections. Nevertheless, we formulate the result already here.
Theorem of the visual body model for compound eyes with convex curvature
If a segmented bilateral living being has compound eyes with convex curvature so that an upright and laterally correct image is formed on the approximately round visual receptor surface, the receptor axons (directly or after switching to projecting ganglion cells) move to the assigned visual segment and contact visual connective neurons there. These form a body image of the visual receptor surface on the outer surface of a half cylinder. This body image lies above the sensory body images already present there, which form nested mantle surfaces of half cylinders arranged according to modalities. The retinal interior is assigned to the tail-side body segments, the exterior to the head-side segments. There is no rotation or mirroring of the images. The retinal signals of the visual objects on the left activate trunk muscles on the left, those on the right activate those on the right. The top and bottom are also retained within the projection.
The axons of the visual connective neurons transport the signals (as in the rope ladder system) headlong to the topmost turning floor. There, signal reversal takes place via commissure neurons, which are synaptically connected to descending projecting motor connective neurons. The latter project to the motor neurons so that visual signals generate movements directed towards the visual object.
Such a projection of an approximately circular receptor surface onto the mantle surface of a half cylinder is called an unreflected projection.
Theorem of standard visual motor projection with body side mirroring in lens eyes
In lens eyes, there is a standard visual motoric projection with body side mirroring.
Pit eyes, which work like a pinhole camera, are equal to lens eyes in their mode of operation.
This leads to the assumption that the projection axons of the trunk senses, which reach the head area in the segment of lens eyes, must first be subjected to a body side mirroring in order to ensure that optical stimuli cause movement in the direction of the visual object. The descending, motor output must also be subjected to a body side reflection after leaving the optical segment in order to control the muscles that cause movement towards the optical stimulus object.
A body side mirroring can be achieved by crossing the axons of the left side of the body to the right side in a certain segment located on the head side, while at the same time the axons of the left side switch to the right side. Such axon crossings have been demonstrated in vertebrates and many theories have tried to explain the formation of such crossings. We postulate here the formation of such a signal crossing in the fifth segment and will refer to this as crossing segment.
This raises the question as to what cause led to the formation of such a sensory and motor lateral body mirroring. The answer is provided by the study of the primordial vestibular sense.
The imaging topology of the primordial vestibular sense
We are now analysing the imaging topology of the primordial vestibular sense in which a rolling grain of sand in a statocyst irritated the hair cells at the deepest point. If we project the spherical inner surface of the statocyste and the hair cells contained in it onto a circular surface, a vestibular circular function iscreated.
First, we transfer this into a vestibular semicircular function, as was already done with the visual signals. We can map this by a simple topological mapping to a vestibular mantle function defined on the mantle surface of a half cylinder. We imagine the vestibular receiving neurons arranged on the mantle surface, which receive the vestibular signals via axons with the hair cells (possibly with the interposition of auxiliary neurons). Each receiving neuron takes over the fire rate of its receptor.
However, here we are faced with a problem: An active hair cell must activate a trunk muscle on the contralateral side. This is explained using the example of a human being.
If a person bends forward, the abdominal trunk muscles are activated and cause the bending. A grain of sand in the statocyste would roll forward. However, in order to return to an upright body position, the excited hair cell at the front must activate the trunk muscles at the back so that a counter movement is initiated into the vertical.
Each hair cell must therefore be connected to a contralateral motoneuron so that a corrective movement restores the standard position.
The whole thing is complicated by the fact that the sensory signals - which also include the vestibular signals - can only run along the sensory half in the neural tube, because the other half is assigned to the motor signals and their axons. Nevertheless, signal crossing to the contralateral side is also possible here.
We will now compare the uncrossed and crossed signals, separated according to sensory and motor signals. For this purpose, we think of the neural tube being cut in the vestibular segment so that the cut surface is circular.
In the following figure we divide the neural tube into a sensory part (yellow) and a motor part (green). We also divide it into four right sub-segments (1R to 4R) and four left sub-segments (1L to 4L).
Shown is the arrangement of the neurons of the neural tube in the trunk area and in the head area, between both we think of the crossing floor. The consequences after passing through the crossing floor are shown in the following figure. The ventricular cavity of the neural tube has been omitted for simplicity.
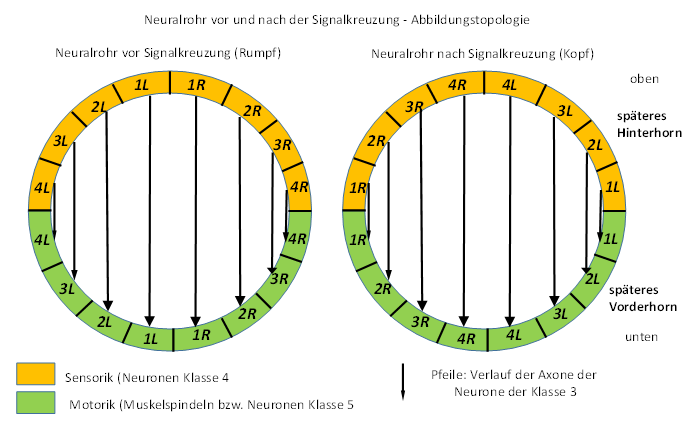
Figure 9 - Principle of signal crossing on the crossing floor
If you look more closely at the above figure, you can see that the quarter circle formed by the sensory neurons of the trunk, which were labeled 1L, 2L, 3L and 4L, simply needs to be rotated 90 degrees to coincide with the neurons 1L, 2L, 3L and 4L of the head, with the center of rotation being the neural tube center.
Similarly, but in the opposite direction of rotation, the quarter circle of the right trunk neurons must be rotated.
At motor neural-tube-half analog each quarter must be turned around centre by 90 degrees, the other half by -90 degrees.
This rotational transformation does not result in any mixing of the axon lines, even the axons of the commissure class 3 neurons, shown as arrows in the figure, run completely ordered and parallel to each other after the signal crossing as before the mirroring.
A complete crossing consists of two partial components: a sensory crossing
and a motor crossing. In the sensory crossing, the ascending sensory signals
move to the opposite side via the crossing axons.
In motor intersection, the descending motor signals travel via axons from the
opposite side back to the side that delivered the ascending sensory signals to
them. These crossings still exist today in the
vertebrate brain.
The sensory signals rise in the Fasciculus gracilis and Fasciculus cuneatus uncrossed and unconnected to the sensory nuclei of the entrance floor, the Nucleus gracilis and the Nucleus cuneatus. Their axons cross as fibrae arcuatae internae of the Lemniscus mediales in the decussatio lemnisci medialis to the contralateral side and from there they move further headwards to the thalamic floor. It can be assumed that this axon course, which is present in humans, manifested itself in the primal brain of chordates in earliest prehistoric times. For the ascending sensory signals, the change to the contralateral side occurred after passing through the sensory input nuclei of the primal brain.
The decussatio tegmentalis anterior is located approximately at the level of the superior colliculi. It contains nerve fibres of the nucleus ruber, which cross to the other side to γ-Motoneurons in the anterior horn of the spinal cord.
The descending motor signals reach the nucleus ruber. From there, the axons move to the contralateral side to reach the ϒ-Motorneurons to descend. These crossing axons form the decussatio tegmentalis anterior. We can certainly transfer these conditions present in humans to the primal brain of the chordata, since axon progressions have a very conservative character.
Theorem of the crossing of afferent ipsi- and contralateral axons in the crossing segment above the input segment
On the head side above the entrance segment there is a sensory crossing segment. The afferent, sensory connective axons of the left and right posterior cortex exchange positions in the crossing segment.
Below the input segment is the motorized crossing segment. The efferent, motor connective axons of the left and right anterior horn exchange their positions in the motor crossing segment. This results in contralateral body imaging in the sensory and motor body models located above the head.
The side change takes place in the ascending part above the input cores and in the descending part below the output cores.
This signal crossing at a very early evolutionary stage has been preserved in vertebrates until today.
Theorem of the visual body model for lens eyes
In lens eyes, the visual image on the retina is displayed upside down and laterally reversed. This corresponds to a point mirroring. This must be corrected by a point mirroring in the axon course. This is done in the intersection of the fifth segment. This is the only way to ensure that the muscles of the trunk of an object in the field of view are supplied with contraction signals which lead to a movement towards this object (foraging).
Theorem of commissure connections between sensory and motor body images in the trunk area
The neurons that represent the sensory body image for a particular modality in a particular segment project via commissures to the corresponding motor body image. Both are located on the lateral surface of one half cylinder each. Both half cylinders form a whole cylinder. In this cylinder, the commissures would all run parallel to each other horizontally from the sensory to the motor half cylinder if the ventricle cavity in the neural tube did not exist and the number of sensory neurons coincided with the number of motor neurons. The parting plane between both half cylinders would then be a symmetry plane for all commissuraxons. However, due to the existing ventricular cavity, the real axon course is curved, and thus no mixing of the axons occurs.
If the number of sensory neurons is much greater than the number of motor neurons, there is a convergence of sensory projections on the motor neurons, which are present in smaller numbers. However, there is always a signal flow from the sensory to the motor body image.
Comment:
At this point it becomes very clear how necessary a strict separation of the sensory from the motor part is. If the ascending, sensory axons were wildly mixed up with the descending, motor axons, there would be no such topological relationships and thus no organized algorithm for systematically influencing motor functions via the sensory part.
Hint:
The cylinder containing the sensory and motor body image of a particular segment need not be nearly as high as the corresponding body segment itself. For example, if the body consists of 20 segments, of which each segment contains only 12 trunk muscles, the sensory body image formed to the muscle spindles (or their analogously acting free nerve endings of neurons) consists of 240 connecting neurons in the uppermost segment, which in turn supply 240 horizontally running commissure neurons with input. The mantle surface of the sensory half-cylinder formed by them is relatively small, because the distance between two neurons on this surface is less than a tenth of a millimeter. The same applies to the motor body image, in which 240 neurons project to the motor neurons of the trunk. The body image thus occupies only a fraction of the available space in the neural tube segment. Only when the number of receptors of certain modalities (such as the sense of touch) increases sharply does the body image grow, and the corresponding body images of other modalities grow simultaneously, even if they have significantly fewer receptors. Only in this way can neighborhood relationships of the body be preserved in the body images of the different modalities.
Theorem of vestibular projection
The ascending and descending trunk signals are subjected to contralateral endoscopy in the crossing floor before reaching the vestibular segment. Therefore, the vestibular signals can be directly transferred into a vestibular mantle function. The vestibular mantle function acts as the mantle surface of a half cylinder and is attached to the existing sensory half cylinders without the need for prior signal mirroring.
In all segments above the intersection segment of the segmented bilateria described here, the sensory and motor body models were contralateral mirrored. Thus, the primary visual segment also received the contralateral mirrored sensory body images of the trunk and its output was projected into motor neurons of the opposite side. Therefore, the concave compound eyes there were no longer useful for targeting dark visual objects. Here the gradual conversion of these eyes in the direction of lens eyes began.
Also in the vestibular segment there was a mean value core that integrated the vestibular signals. This nucleus was the Nucleus Deiters - this is what it is called in mammals. It was supposed to take over a special task in rebuilding the vestibular system.
Theorem of the Nucleus Deiter
The nucleus deiter was (originally) the mean nucleus of the vestibular segment.
Theorem of conversion of primary convex compound eyes into lens eyes
The output of the primary, convex compound eyes of the second segment was no longer useful for a meaningful body control after the development of the vestibularly based contralateral signal reflection in the crossing floor, because instead of the muscles working towards the target, their opponents were now controlled. Therefore, a gradual reduction of these ocular systems was carried out. As a result, the originally convex curved surface became inwardly sinking and concave over the course of countless generations. This restored the usefulness of the visual signals, because this eye shape needed crossed signal paths. Therefore, the concavity was increased, resulting in pit eyes. To prevent foreign bodies from entering the eye pit, a translucent cover was formed over the cavity from which the eye lens developed. Filling the cavity with water and later with translucent tissue served to maintain its shape.
Theorem of the retention of secondary compound eyes
The secondary eyes, which were located below the crossing segment, remained convex compound eyes, because they needed an uncrossed connection which was present there. However, because the imaging quality of the lens eyes that had been formed in the meantime was enormously improved in the course of evolution, the secondary eyes only had a strongly reduced visual function. They specialised in the recognition of light and dark and used these mean value signals to control the living being circadianly by supplying the nucleus suprachiasmaticus on the subthalamic level with synchronising signals.
The two secondary eyes gave rise to two structures in vertebrates. One of these eyes could develop into the parietal eye (parietal eye), which is present in original vertebrates and serves to perceive differences in brightness. The other secondary eye could transform into the pineal gland, which serves the circadian rhythm in the epithalamus.
Theorem of the fusion of the head segments
The head segments specialized in the evaluation of the signals of the dependent receptors. At the same time, these segments merged and were now located in front of the actual mouth opening. Parallel to this, on each side of the body of the head region, both the sensory centers and the motor centers of the head region merged with the exception of the first (olfactory) segment. As a result of this fusion, the respective sensory and motor centers of the fused segments were no longer located one behind the other in the associated rope ladder model, but next to each other.
We interpret the merging of both the head-side segments and the associated sensory and motor centers of the involved segments as the standard case, which was also performed in those segmented bilateria that led to the vertebrates.
In the segmented Bilateria living today, the merged sensory and motor centers of the head segments formed the upper throat ganglion. Below the mouth opening there is often an undermouth ganglion.
Theorem of equal rank of head modalities
After the fusion of the head segments and the fusion of its sensory and motor centers, the head modalities were of equal importance. In the rope ladder model this meant that there was a torso ladder on each side of the body. However, this ladder branched at its uppermost rung into as many ladder sections as there were head modalities, and there were also additional ladder sections for the different torso modalities. For each head and trunk modality there was a separate cranial modality ladder with two rungs. The lower rungs of the different head modalities lay parallel to each other and formed the thalamic center, which is called the upper throat ganglion in today's Bilateria. We call this structure the thalamic turning floor. The sensory or motor centers formed the sensory or motor thalamus, which was divided into the different modalities. The upper rungs of the head are called cortical turning floorsor cortical turning loopsof the respective modalities.
Initially, there was a division into three cranial modality ladders and thus into three cortical turning levels, which are referred to here as temporal, parietal and occipital turning levels. In mammals, the lobi of the brain, which are called temporal lobe, parietal lobe and occipital lobe,originated from them.
Below the thalamic turning floor are the input floors for the head and trunk senses. The visual signals arrive directly at the level of the thalamic turning floor, the olfactory signals even directly at the cortical floor of the temporal loop.
The unfolding of the rope ladder system in modality ladders is sketched in the following figure, where
TS Temporal Loop
PS Parietal Loop
- OS Occipital Loop
On the left is the early state, on the right the later state.
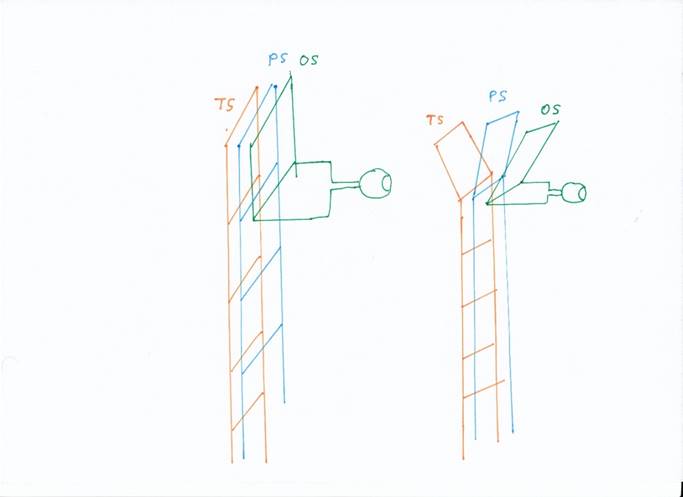
Figure 10 - Splitting the rope ladder system into
modality ladders
In early prehistoric times there was the following distribution of the modalities to the three modality loops:
- The temporal loop mainly received signals from the hair cell receptors. These served on the one hand the vestibular system, on the other hand the lateral line sense of the trunk. With the further development of the vestibular system, the sense of hearing emerged, whose signals were also evaluated in the temporal loop.
Furthermore, the temporal loop received the olfactory signals directly - i.e. without a detour via the thalamic level - and later also included the limbic system that was forming.
- The parietal loop received signals from the trunk's muscle tension receptors and tendon organs as well as signals from other trunk receptors that analyse joint positions. With the development of sensors for the sense of touch and pain, their signals were also evaluated in the parietal loop. Later, the analogue signals of the developing fins or the extremities of the tetrapods were added.
- The occipital loop received the output of the visual receptors.
A special feature of the temporal loop was that it contained the olfactory system as a subsystem, which was later extended to the limbic system. In particular, the sensory input core and the motor output core of the olfactory floor became independent structural elements of the temporal loop or the resulting temporal lobe of vertebrates. The same applies to the mean nuclei of the olfactory floor. One became a subcore of the amygdala due to its spatial proximity. Even the projection of the olfactory level into the dopaminergic mean nucleus ended in a separate structure called area tegmentalis ventralis, or VTA for short. And while the three (later four) cortical modality loops and the resulting lobi used the nucleus basalis Meynert as the cholinergic mean nucleus, the septum within the temporal loop performed the function of the cholinergic mean nucleus belonging to the olfactory level.
Even the striatum, the origin of which will be described later, had a counterpart in the olfactory level and, also due to its spatial proximity, became a subcore of the amygdala, characterized by GABAergic neurons with dopaminergic entry and D1 or D2 receptors for dopamine.
Another cortical turning loop was created when a new neuronal structure called the cerebellum developed. The output of this new structure led to the formation of the frontal turning loop, from which the frontal lobe of vertebrates emerged. This will be described later.
Theorem of the unfolding of the cortical floor in modality loops
The unfolding of the cortical turning loop into modality loops led to the formation of the temporal loop, the parietal loop and the occipital loop, each of them processing signals of different modalities.
With the much later development of a cerebellum, the frontal turning loop continued to develop.
From these modality loops, the temporal cortex, parietal cortex, occipital cortex and frontal cortex developed in vertebrates.
The unfolding of the modality loops had consequences for the cores present on this floor. While in the temporal loop, the cholinergic mean nucleus was formed by the septum, in the other loops the cholinergic mean neurons joined together to form a structure known as Nucleus basalis Meynert.
The modality loops resulting from the splitting of the modalities also possessed a topological order that can be derived from the previously existing topological properties.
The class 4 neurons, which were in the cortical turning loops, could no longer project to higher levels of the nervous system because the cortex formed the highest level. Therefore, they formed into local interneurons that received the ascending input and passed it on to the existing class 2, 3 and 6 projection neurons. Since they were originally only present in the sensory centers, they only existed in the sensory cortex of the different Lobi.
Theorem of sensory body images in the cortex
The input neurons of class 4 of the sensory cortex surface, which may be represented simplified per half of the body on the surface of a half cylinder, form a vertical strip for each sensory modality. Each modality strip consists of horizontally arranged segmental strips. The topmost, first segment strip is assigned to the tail segment. Thus, in each modality strip of the cortex surface, a body model is created by the input neurons, in which for each modality the distribution of the corresponding receptors of this modality on or below the surface of the body of the living being is represented.
In the cortex surface, the input neurons of class 4 form a single layer.
A transformation from half cylinder to hemisphere, which better approximates the shape of a hemisphere in mammals, results in analogous modality and segmental stripes, but these are now projected onto the shape of the sphere.
The unfolding of the modalities in the thalamic floor leads to the cortex area initially consisting of three, later four lobes, which is responsible for related groups of receptors.
The axons that are assigned to the tail segment run inside the neural tube, while the axons of the higher segments are located outside. When they reach the cortex, the axons of the uppermost segment are located on the outside, whereas those of the tail segment are located on the inside. In this order, the axons of the ascending connective neurons dock to the cortex neurons of layer 4. Therefore, the tail segment is located at the top of the cortex in this projection, the order of the segments is quasi "upside down".
It should be recalled that the sensory cortex emerged from the sensory centre of the first segment. This projected in the rope ladder nervous system of each half of the body to the motor centre of the same segment. This projection was preserved and led - also by unfolding - to the development of a motor cortex.
A local increase or decrease in the number of receptors of a modality, which develops in the course of evolution, leads to spatial distortions, and the associated surfaces in the cortical body model then take up relatively more or less space. Therefore, very sensitive regions are shown enlarged, while receptor poor regions are reduced.
With the development of new body parts such as head muscles, jaws, fins or later, in the case of the tetrapods of limbs, new model parts are inserted into the existing body models, the position of which depends on where in the thalamus the ascending sensory input arrives.
The input reaches the neuron layer of the class 4 neurons in the sensory cortex, we briefly recall the rope ladder nervous system and the tetraneural system. There, the input arriving in the segment was passed from the class 4 neurons via class 3 connective neurons to the class 5 motor projection neurons and from there sent tailwards to its associated motor neurons.
Likewise, the input of neuron class 4 was sent via cross-commissures of neuron class 2 to the opposite side, i.e. the opposite half of the cortex, for the purpose of contralateral inhibition. These cross-commissures were present in both the sensory and motor cortex. The associated projection axons form the bar (corpus callosum) in humans.
Likewise, the cortex surfaces contain neuron class 6 of the mean value neurons and neuron class 1 of the activation neurons, which are connected to the mean value centers.
In the motor cortex we find the neuron class 5 of the neurons projecting to the neural tube or spinal cord.
Thus we find all six neuron classes in the uppermost cortical level. Sensory neurons are located in the sensory half of the cortical floor, the motor neurons in the motor half.
We assume that the class 6 neuron that projects to the respective mean centre of a segment was the first to form in the course of evolution, because viability was the first to be ensured. Consequently, the projections from the sensory to the motor centre of a segment emerged afterwards, even before the formation of segmented bodies. Thus, neuron class 3 developed after neuron class 6, and at about the same time the projection from a centre to the contralateral centre via neuron class 2 developed.
According to this, with the formation of segmentation, the sensory and motor connective neurons must have been developed, which enabled the exchange of signals between the segments. These are the neuron classes 4 and 5, which were moved between class 6 and class 3. The class of activation neurons of class 1 was the last to develop, which attached itself externally. Of course, this hypothesis only applies to segmented bilateria whose line led to the vertebrates. We can thus formulate another theorem for these organisms.
Theorem of the well-being order of the different neuron classes in the neural tube
In the neural tube of the living beings whose line led to the vertebrates, a well order of the different neuron classes developed in independent layers from the outside to the inside in the following order:
Layer 6 of neuron class 6 (mean neurons)
Class 5 motor commissure neurons layer 5
Layer 4 of sensory commissure neurons in class 4
Layer 3 of sensory connective neurons in class 3
Layer 2 of class 2 motor connective neurons
Layer 1 of class 1 activation neurons
Here, layer 1 with the class 1 neurons is located directly adjacent to the ventricle space of the neural tube. The proximity of the ventricle space to the class 1 neurons seems to be essential for the stratification direction.
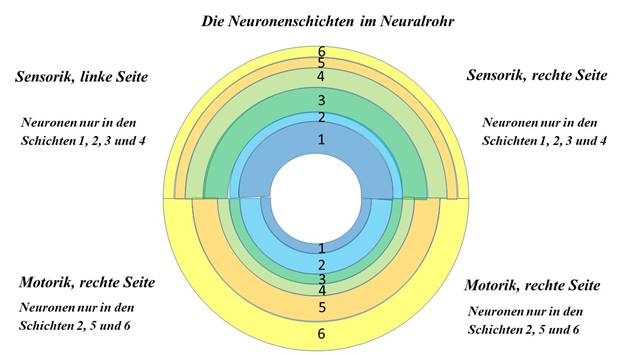
Figure 11- Arrangement of neuron classes in the neural tube
We now analyze the topology of the class 5 neurons in the first segment and remember the rope ladder model again. We align it so that the head is at the top. The class 5 neurons are located in the motor center, which receives its input from the sensory center. This, however, was split on the first level according to the different modalities and now consisted of adjacent modal strips of class 4 neurons, which, according to the rope ladder model, transmitted their excitation (via interneurons) to the projection neurons of class 3, whose axons now moved to the motor neurons of class 5.
The projection axons of these neurons fanned out in the same way as those of the upwardly projecting class 4 neurons, but they consisted of only one type of modality. Each class 5 neuron projected to motoneurons - so there was only one modality. So the area where the class 5 neurons were located formed a contiguous modality area, which we call the motor cortex.
However, the cortical modality area was organized in stripesjust like the sensory modality areas. Each body segment was assigned exactly one horizontal strip. The uppermost strip represented the lowest segment. Each higher segment projected into a lower segment strip. Thus the motor cortex was also a segmented body model of the segmented living being. In this model, the motor neurons were represented by the output neurons of class 5. We summarize this finding in a theorem.
Theorem of motor body imaging
The motor cortex represents a body model. In this model, the output neurons of class 5 represented the real motor neurons. Each segment occupied a separate segment strip in the model. Neighborhood relationships of the motor neurons were represented in the body model by the corresponding output neurons.
We assume that the sensory and motor cortex of animals of the evolutionary epochs of that time retains the stratification in neuron classes.
Theorem of the stratification of neurons in the cortex according to neuron classes
In the cortex of the segmented Bilateria, whose line led to the vertebrates, the neurons in the cortex - which emerged from the first segment of the early segmented Bilateria - were stratified according to neuron classes. The layering is exactly the opposite of the neural tube. From inside to outside, the following layers were formed:
Layer 6 of neuron class 6 (mean neurons)
Layer 5 of motor connective neurons of class 5
Layer 4 of sensory connective neurons in class 4
Layer 3 of sensory commissure neurons in class 3
stratum 2 of motor cross-commissory neurons in Class 2
Layer 1 of class 1 activation neurons.
The layering sequence in the cortex is the opposite of the layering direction in the neural tube. This anomaly is caused by the close coupling between the first layer and the ventricular space. In the neural tube the inner ventricle space serves as a reference. In the cortex, the distance to the inner ventricle space is much too great for coupling. Here the outer ventricle space takes over this function, therefore the order of the neuron layers is reversed. The layer with the class 1 neurons is located outside directly on the outer ventricle wall.
Layer 4 of the sensory commissure neurons in the cortex - i.e. the highest segment - has a special feature. In the remaining segments, these neurons are projection neurons, which project into the superior segment. In the highest segment, however, there is no superior segment. Therefore, these neurons regress to interneurons, which distribute the received excitation to the neighbouring layers.
Not every layer contained neurons everywhere. On the sensory half were the sensory neurons, on the motor half the motor neurons.
In the motor cortex there were no class 4 sensory commissure neurons and no class 3 sensory connective neurons, because these were already in the original rope ladder system on the sensory side of the neural rope ladder. Class 2 neurons were present, as were class 1 and 6 neurons.
In the motor cortex, only neurons could be present that were also present in the motor centre at the motor spar of the rope ladder system.
Likewise, in the cortical sensory strips, which in their entirety formed the sensory cortex, there were no motor commissure neurons of class 5, but instead there were the sensory commissure neurons of class 4 (as interneurons) and the sensory connective neurons of class 3. neurons of class 2 were present, as were neurons of classes 1 and 6.
In the sensory cortex, only neurons could be present that were also present in the sensory center at the sensory spar of the rope ladder system.
Already quite early in the evolution of Bilateria, the first components of a motor contralateral inhibition developed. If a worm-like creature had a ring muscle tube and a longitudinal muscle tube, these worked against each other. The contraction of a ring muscle was only successful if the longitudinal muscle did not oppose it. The contraction of the longitudinal muscles required a decontraction of the ring muscle. This could be achieved if both types of muscle inhibited each other so that the more active muscle prevailed. In the case of the bilateria, this mutual inhibition was extended to contralateral inhibition. While motor contralateral inhibition initially developed in the neural tube or spinal cord and was later transferred to the brainstem, sensory contralateral inhibition couldonly establish itself in the cortex, because only there were all sensory signals of the trunk and head senses available. Sensory inhibition was not about one muscle asserting itself against its opponent, but rather about the stronger sensory signals of one hemisphere of the brain asserting itself against the weaker signals of the other hemisphere. This was realized separately according to modalities. The neural circuitry required for this is described in the following theorem.
Theorem of contralateral (sensory) inhibition in the interaction of thalamus and cortex
The thalamic connective neurons of class 4 of one half of the body project to the equilateral cortex into the neurons of layer 4, which transmit these excitations to the commissive neurons of class 3. The latter project to the motor cortex.
However, the excitation of class 4 neurons was also transferred to class 2 neurons, which projected to the opposite side into the sensory contralateral cortex half. There they contacted class 5 neurons whose axons descended to the thalamus of this half of the body, where they caused contralateral inhibition via inhibitory neurons. The inhibitory neurons formed the nucleus reticularis thalami.
Thus, there was a cortical upward projection in the thalamus and a cortical downward projection of the same thalamic neuron in the immediate vicinity with the interposition of an inhibitory neuron. However, not only the cortex cortex projects back to the same thalamic neuron, but the downward projection originates from the mirror imaged contralateral cortex cortex cortex and causes the contralateral inhibition in the brain system.
This circuit is realized in both halves of the brain. Point by point, receptor by receptor, the signals of the left and right hemispheres of the body are compared and inhibit each other. Only the stronger signal with the higher rate of fire will prevail and can cause reactions in the system. The cortical back projections to the thalamus originate from the opposite side and realize the contralateral inhibition on the level of the second segment.
In the vertebrate brain, the myelinated axons of the cross-connective of the first (cortical) segment that conducted the signals to the contralateral side form dense axonal bundles called bars or corpus callosum.
The thalamic inhibitory neurons formed a layer of neurons outside the thalamus, which received its input (mainly) from the first segment - the cortex - and had an inhibitory effect on the associated thalamic neurons. This layer also belongs to the thalamus and is called the reticular nucleus thalami. Since it receives its input from the class 5 neurons, it is only present on the motor side of the thalamus.
Surgical severing of the corpus callosum, known as split brain, which was previously used in epilepsy, for example, leads to the elimination of neuronal competition between the two hemispheres of the brain, so that in extreme cases an independent sensory awareness can develop in each hemisphere of the brain. It is then as if each half of the body were an independent living being. The motor function is less affected by this, because the neuronal competition of the motor function continues, on the one hand on the different segmental levels of the trunk and on the other hand via the cerebellum, which replaced the contralateral inhibition on the brainstem level by the inverse excitation of the motor opponents.
As a result of the division of labour between the halves of the brain in Homo sapiens, each of the two levels of consciousness after a brain-split operation only has the abilities that are localised in the respective half of the brain.
A disturbance of the signal conduction in the corpus callosum or in the nucleus reticularis thalami could lead to the effect of a split of consciousness in which the person concerned is represented by two personality variants (schizophrenia).
Of course, the question arises why the contralateral inhibition of the two cerebral hemispheres was not realized by inhibitory interneurons of the cortical floor.
Theorem of contralateral inhibition of the two hemispheres of the brain
The contralateral inhibition between the two cerebral hemispheres was realized by inhibitory interneurons of the nucleus reticularis thalami because the cortical floor had already transferred its inhibitory interneurons to the striatum. Therefore, this inhibition had to be realized one floor below by thalamic neurons. During evolution, these neurons formed a separate thalamic nucleus, the nucleus reticularis thalami.
The use of the inhibitory interneurons of the cortex for the forming striatum will be discussed in more detail in the chapter on the basal ganglia. This is one of the few cases in this monograph where knowledge is used before it is derived.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan