Theory of the brain’s neural circuitry
and analytical thinking
ISBN 978-3-00-037458-6 - ISBN 978-3-00-042153-2
Part 1.6 The neural elementary oscillating
circuits in the striosome system
In sketch 1.9 the globus pallidus interna was shown as a negative negation nucleus for the striosome signals.
In the diagram, for each activity neuron there is an exciting connection to a dopaminergic neuron of the substantia nigra pars compacta. This neuron excitably projects onto a GABAergic striosome neuron whose axon branches out. One axon branch supplies the globe pallidus interna, while the second branch projects back to the neuron in the substantia nigra pars compacta.
The GABAergic projection of the striatum onto the substantia nigra pars compacta is so far (in July 2011) theoretically unexplained.
This monograph shows for the first time a meaningful interpretation of the task of this projection.
This becomes all the more important because the origin of the climbing fibre signal and its function can now be scientifically interpreted and proven.
It is known from physics that a signal that is generated in a system can be traced back to its place of origin. In this case one speaks of feedback.
In electrical oscillating circuits, oscillations are thus created which would not be possible without feedback.
Therefore, the author postulates the emergence of a vibration here as well. This oscillation may be interpreted as the system clock of the corresponding activity neuron. If the origin of the oscillation can be proven, a cortex cluster could be interpreted as an elementary computer of the brain with its own system clock. The striosome system would be a multiple parallel connection of such elementary computers.
Theorem 1.13: Elementary oscillating circuits in the striosome system
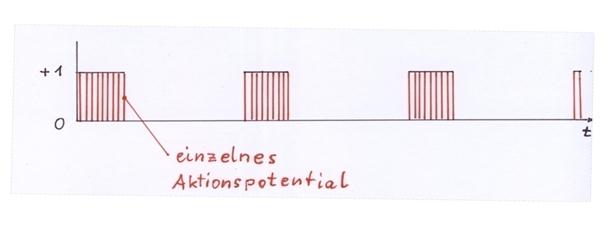
One activity neuron of each cortex cluster, the dopaminergic neuron in the substantia nigra pars compacta excited by it and the GABAergic neuron of the striosomes of the striatum excited by the latter neuron form a neuronal elementary oscillating circuit as neuron triplets. This neuronal elementary oscillating circuit receives as input the mean value signal of a cortex cluster. In the case that this mean value signal is not the zero signal and lasts for a longer period of time, the neural elementary oscillatory circuit transforms this temporary continuous signal into a system clock, which constantly changes between the signal values 0 and 1 as an envelope signal. The envelope frequency of this neural system clock is about 5 Hz, so that about 5 oscillations per second occur, while the rate of fire below the envelope is quite high.
Each cortex cluster generates its own system clock in case of the activity of the corresponding activity neuron. Nevertheless, the system clocks of the different, neighbouring clusters are loosely coupled with each other, which enables a temporary synchronous oscillation.
It will be shown later that the neural system clock of a cluster is used to generate a climbing fiber signal, which is used to store the signal combinations occurring in this cluster.
Proof of theorem 1.13:
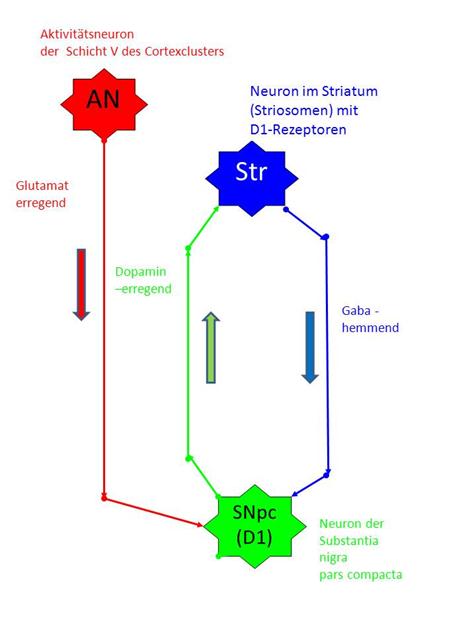
We record an elementary oscillating circuit of the striosome system consisting of three neurons, taking into account the spatial arrangement of these neuron types in the brain.
At the top, the activity neuron AN of layer V of the corresponding cortex cluster is shown in red.
Below is the neuron SNpc of the substantia nigra pars compacta of the D1 system, shown in green.
In between is the associated GABAergic neuron STR of the striatum (blue).
It is internationally undisputed that the neurons of the striosome system of the striatum of the basal ganglia project into neurons of the substantia nigra pars compacta by means of the Gaba transmitter. There are two classes of neurons in the substantia nigra pars com-pacta. The first class, to which our neuron SNpc should belong, projects into the striosomal neurons using dopamine, whereby the receptor type D1 has a dopamine-exciting effect (subtype D1).
Sketch 1.18: Elementary oscillating circuits in the striosome system
The second class of neurons of the substantia nigra
pars compacta, which is initially neglected here, projects into the matrix of
the striatum by means of dopamine and there has an inhibitory effect on the
target neurons (subtype D2) via D2 receptors.
We draw the three corresponding axons in the above circuit diagram of the neural elementary resonant circuit:
· An excitatory glutamatergic axon leads from the neuron AN of the cortex cluster to the neuron SNpc of the substantia nigra pars compacta of type D1 (red).
· From the neuron SNpc of the substantia nigra pars compacta subsystem D1 an excitatory, dopaminergic axon leads to the neuron Str of the striosomes (green).
· An inhibitory GABAergic axon leads from the neuron Str of the striosomes to the neuron SNpc of the substantia nigra pars compacta subsystem D1 (blue).
It is undisputed that dopaminergic projection uses weakly myelinated axons, which means that the action potentials located on them spread more slowly.
We prove the origin of the neuronal system clock, which may be the envelope of a sequence of action potentials, under the following - not necessary - conditions:
1. The distance between the STR neuron of the striosomes and the SNpc neuron of the substantia nigra pars compacta is (as an example) 50 mm.
2. The frequency of the activity neuron AN is (exemplarily) 200 Hz, so that it fires 200 action potentials per second.
3. The speed of the action potentials along the axons is (as an example) 1 meter per second, i.e. 1 mm per millisecond.
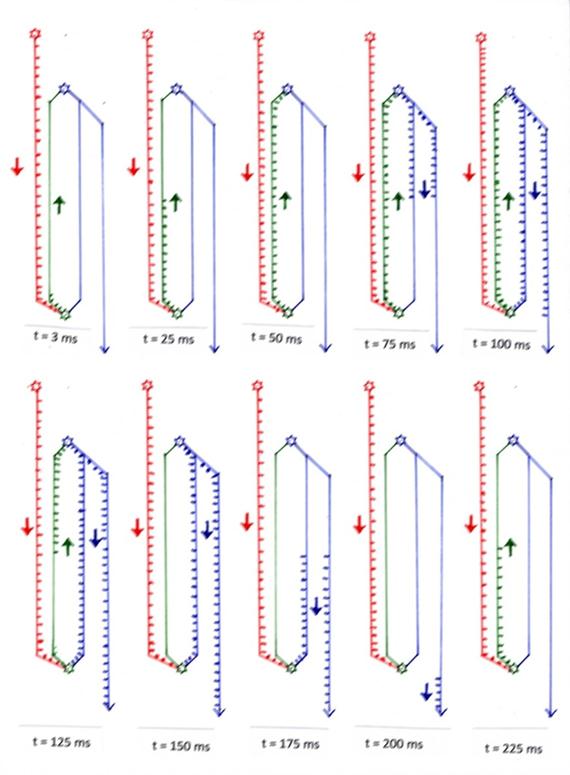
At time t = 0 ms, the two neurons STR and SNpc are inactive. The continuous signal from activity neuron AN already arrives at the neuron SNpc. Each incoming action potential of the AN neuron (red) causes the creation of an action potential at the SNpc neuron. These dopaminergic action potentials, which are generated there, spread along the axon shown in green towards the STR neuron. After t = 3 ms they have covered a distance of 3 mm, as can be seen in the first image of the slow-motion display. At the time t = 25 ms, these action potentials, which are still continuously being generated, have covered half of the distance, i.e. 25 mm (Figure 2).
After 50 milliseconds (Figure 3), the dopaminergic action potentials finally arrive at the STR neuron of the striosomes for the first time. There, due to their excitatory effect, they generate action potentials, which now move in the opposite direction to the SNpc neuron. After 75 milliseconds, this gabaergic action potential has already completed half of the way back. At the time t = 100 ms the GABA-ergic, i.e. inhibitory action potentials reach the SNpc neuron for the first time and cause a strong inhibition there, which may last at least for the short time of 5 milliseconds (refractory time).
However, this means that the previous dopaminergic action potentials do not occur, since the associated SNpc neuron is now permanently inhibited as long as inhibitory action potentials reach it. At time t = 125 ms, the SNpc neuron is already silent for 25 milliseconds, while action potentials are still available for the inhibition period of 75 milliseconds on the oscillatory circuit axons (blue and green).
After 150 milliseconds all action potentials of the dopaminergic axon are used up, but the gabaergic axon still contains inhibitory action potentials for exactly 50 milliseconds. At time t = 125 ms, these are only half used up.
Exactly at the time t = 200 ms the last inhibitory gabaergic action potential arrives at the SNpc neuron and inhibits it for the last time. After this time, all previous action potentials of the blue-green pathway are used up, and a possibly still existing excitation of the cortical activity neuron AN can again lead unhindered to excitation of the SNpc neuron. The process starts again.
To sum up, it can be said that
1. For a period of 100 ms the neuron of the substantia nigra pars compacta is excited and sends out action potentials.
2. For a period of 100 ms the neuron of the substantia nigra pars compacta is inhibited and does not send out action potentials.
3. For a period of 100 ms the neuron of the striosomes of the striatum is excited and sends out action potentials.
4. For a period of 100 ms the neuron of the striosomes of the striatum is inhibited and sends out action potentials.
5. There is a phase shift (time shift) of 100 ms between SNpc neuron and STR neuron with regard to excitation and inhibition.
6. In the shown system of three neurons there is a neuronal oscillation which has a frequency of 5 Hz for the given numerical examples, i.e. it has 5 oscillations per second. This oscillation is the envelope curve of the involved action potentials. The action potentials under the envelope have the frequency of 200 Hz, as given at the beginning of the proof (example).
Slow-motion display of the oscillations in the striosome oscillating circuit
Sketch 1.19: Oscillation in the striosome system - the neuronal system clock of a cluster
Theorem 1.14: Loose synchronization of the oscillations in the striosome system
The many individual resonant circuits in System Globus pallidus interna, substantia nigra pars compacta (subclass D1) and in the striosomes are partially synchronized by loose coupling in the substantia nigra pars compacta.
Proof of Theorem 14: Verbatim quotation from "Functional Neuroanatomy" Zilles/Rehkämper, 3rd edition, Springer Verlag Berlin Heidelberg 1998, page 287, first and second paragraph, with the kind permission of the author
(begin quote)
"In the substantia nigra, a very cell dense area, Pars compacta, and an area with low cell packing density, Pars reticularis, is visible. While the pars reticularis of the substantia nigra has GABAergic neurons that project onto the motor thalamus nuclei, the pars compacta contains predominantly large dopaminergic neurons that have a long-same (2 - 5 Hz) pacemaker-like activity and whose axons reach the striatum.
Besides the content of melanin, which gives the substantia nigra its name-determining black colour, dopaminergic neurons have several special features. They exhibit synchronous activity, which is attributed to dendrodendritic synapses between neurons and dendritic release of dopamine. The release of dopamine at the axon terminals is regulated more by presynaptic receptors than by pacemaker activity.
(end of quote)
At this point, we would like to thank the editors, Professor Karl Zilles and Professor Gerd Rehkämper for their extremely precise presentation of cytoarchitectonics, especially of the basal ganglia, as well as Professor Rolf Kötter. In particular, the systematic descriptions of the cytoarchitectonics of the Basal Ganglia by Professor Rolf Kötter, which are still available on the Internet today, were the inspiration for the development of this theory.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan