Theory of the brain’s neural circuitry
and analytical thinking
ISBN 978-3-00-037458-6 - ISBN 978-3-00-042153-2
Part 1.4 Inversion of neural signals
Already in the retina there are two types of magnocellular ganglion cells. One type of ganglion cells fires more strongly when the brightness of the corresponding receptors increases. This type of ganglion cells is called the on-type.
A second type of magnocellular ganglion cells reacts in exactly the opposite way. If the brightness at the connected receptors increases, this type of ganglion cell fires more weakly, it is called off-type.
Since the output of these two cell types represents the brightness, the associated signals are called Hell-On and Hell-Off. They could also be called dark off and dark on signals.
It is essential that there are always two ganglion cell types that react inversely to the same signal with increasing signal strength:
- with On-Neuron Non the fire rate fon grows strictly monotonously with increasing signal strength S
- with Off-Neuron Noff the firing rate foff drops strictly monotonously with increasing signal strength S.
We summarize these special features in a separate definition.
Definition 1.15: Inverse neurons, inverse signals
A neuron Non has the fire rate fon = fon(S), where fon in the signal strength interval < Smin, Smax > is a strictly monotonically growing function of the signal strength S.
A neuron Noff has the fire rate foff = foff(S), where foff in the signal strength interval < Smin, Smax > is a strictly monotonically decreasing function of the signal strength S.
If the function fon is symmetrical to the function foff with respect to the interval center SM = (Smax - Smin ) / 2, i.e. the equation
fon( SM + ΔS ) = foff( SM - ΔS )
is fulfilled for all permissible values ΔS, then we call Non the inverse neuron to Noff and Noff the inverse neuron to Non and express this with the following symbolism
Non = N*off
Noff = N*on.
The fire rate fon represents the on-signal to the signal strength S, the fire rate foff represents the off-signal to the signal strength S. This is expressed by the following symbols:
fon = f*off
fon = INV(foff) INV() = Inversion operator
Inverse signals always occur in pairs. Examples are the Hell-On signal and the Hell-Off signal of the magnocellular ganglion cells of the retina.
People tend to give inverse signals their own names, with additional words indicating the degree to which they apply. So there are the terms light and dark. These can be specified by adjectives like strong, something, very, light, etc.: "very light", "very dark", "slightly light", "somewhat dark". This is an indication that a continuous signal strength of an interval is meant here, and the signal values at the inter-values are designated with the terms light or dark. The intensities in between are described by suitable adjectives. Analogously, in visual terms the terms red and green mean the ends of a light frequency interval, but here the intermediate values are not described by adjectives such as strong, weak, medium strong or similar, but the intermediate values are first given their own colour names. Yellow corresponds approximately to the middle between red and green, orange approximately to the middle between red and yellow, while green-yellow in German does not get its own name, but is only assigned as green-yellow approximately the middle of the wavelength between green and yellow. The subsequent color space from yellow to blue is determined by other receptors and leads to the formation of color intervals with their own interval name, e.g. violet. The basis of interval formation in colour vision, but also in many other sensory senses, is the simultaneous coexistence of on and off signals.
From an available original signal, an associated inverse signal can be obtained in a neural circuit. This requires only one neural operation: the relative inhibition of an already existing continuous signal with a relatively constant rate of fire.
Definition 1.16: Relative escapement
A neuron N1 has the firing rate f1 = f1(S), where f1 is a strictly monotonic function of the signal strength S. Its output is the signal S1.
Another neuron N2 generates a continuous signal (single signal) with the rate of fire fd.
A third neuron N3 receives inhibiting input from neuron N1 and exciting input from neuron N2 in such a way that its output S3 is an inverse signal to the signal S1. Then we call the inhibition of neuron N2 by neuron N1 a relative inhibition.
Theorem 1.10: Signal inversion through relative inhibition
An on signal generates an associated off signal by relative inhibition of an on signal. Likewise, an off-signal generates an on-signal by relative inhibition of an on-signal.
There are three types of colour-sensitive receptors in the retina. The original signals are off-signals, as increasing brightness inhibits the excitation of the receptors.
In addition, the retina contains the corresponding on-signals. These arise from the primary off-signals by inhibition of an on-signal. The on-signals are provided by the band synapses in the bipolar cells. Their permanent excitation is inhibited by the off-signals, so that now on-signals are generated.
Since this is true for all colors, there are (theoretically) three types of color-sensitive primary retinal signals:
- Red-on and red-off
- Green-On and Green-Off
- Blue-on and blue-off.
However, these signals do not leave the retina, but are still connected to complex signals in the retina.
This results in a total of five signals, which leave the retina via the associated ganglion cells in the direction of the lateral geniculate corpus:
- S1 = Red-On/Green-Off = rt+/gr-,parvocellular ganglion cell
- S2 = Red-Off/Green-On = rt-/gr+, parvocellular ganglion cell
- S3 = light on/dark off = h+/d-, magnocellular ganglion cell
- S4 = light off/dark on = h-/d+, magnocellular ganglion cell
- S5 = blue-on/yellow-off = bl+/ge-, parvocellular ganglion cell
Here the color signals S1 and S2 are inverse to each other.
The brightness signals S3 and s4 are also inverse to each other.
rt+/gr- = INV(rt-/gr+) = (rt-/gr+)*
h+/d- = INV(h-/d+) = (h-/d+)*.
There is no inverse signal for the signal S5 (in humans and primates). It will be shown that this is not absolutely necessary.
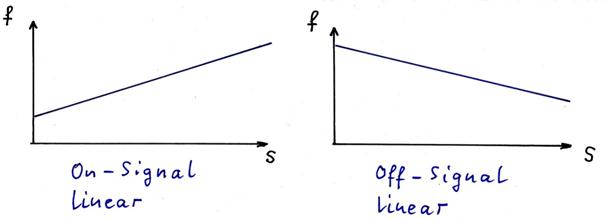
sketch 1.14: inverse signals - special case: linear functions
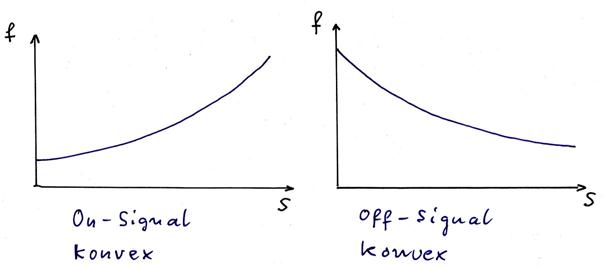
sketch 1.15: inverse signals - special case: convex functions
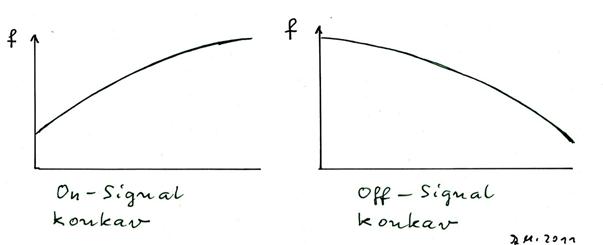
sketch 1.16: inverse signals - special case: concave functions
New signals can often be obtained from two or even three signals. Certain rules can be observed. For example, two or three on-signals are usually combined in an exciting way by a neuron combining their inputs. The signal "Hell-On" is created by excitingly combining the signals "Green-On", "Red-On" and "Blue-On". The signal "Dark-On" is created by combining the signals "Red-Off", "Green-Off" and "Blue-Off", if the brightness still allows color vision.
If, on the other hand, an on-signal and an off-signal are combined with each other, one of the two signals usually excites the resultant neuron, while the second signal inhibits this resultant neuron. For example, the "red-on/green-off" signal is created by applying excitation to the red signal and inhibition to the green signal. Nature seems to prefer the result of a combination of different input signals to be strictly monotonous in the final result. With strictly monotonous functions, there is a maximum of one function value (y-value) for each function argument (x-value), both can be assigned to each other unambiguously.
The choice of strictly monotonous functions prevents ambiguity in the signal value meaning.
Theorem 1.11: Conservation law of strict monotony
If different, strictly monotone signals are fed to a neuron as input, a strictly monotone output signal is usually produced again in case of relative excitation (or) and inhibition.
Conclusions from the conservation law of strict monotony:
If the conservation law of strict monotony applies, there is generally no retinal output of the type
- rt+/gr+ or rt-/gr-
- h+/d+ or h-/h-
- b+/rt+ or b+/gr+
- b/rt- or b/gr-.
Justification: In all of the above cases, a signal function that increases in a strictly monotonous manner with the light frequency is combined with a signal function that decreases in a strictly monotonous manner. The result is a sum function that grows strictly monotonically in one sub-interval, but falls strictly monotonically in a neighbouring interval. This means that a clear assignment of the signal value (the fire frequency) to the current light frequency is no longer so easy, because one signal value would correspond to several light frequencies. Therefore nature prefers those variants where the final result is strictly monotonous to the signal intensity, no matter if growing or falling, the main thing is strictly monotonous. The main purpose is to avoid signal value ambiguities. Due to the irritation that would occur, these would be an evolutionary disadvantage compared to systems where the conservation law of strict monotony is observed.
Definition 1.17: Inversion core with external input signal
A neuron nucleus is an inversion nucleus with an external input signal if it
- receives an exciting input from input signals from a neural region
- receives another inhibitory input from another neural region
- its output neurons each process an input signal and an inhibiting input signal such that the inhibiting signal causes relative inhibition of the input signal, so that the output signal is the inverse of the inhibiting input signal.
Definition 1.18: Inversion core with internal input signal
A neuron nucleus is an inversion nucleus with an internal input signal if it
- receives a stimulating input from a neural region
- forms single signals from this exciting input
- receives a further, inhibiting input from another region
- its output neurons each process a self-generated input signal and an inhibiting input signal in such a way that the inhibiting signal causes relative inhibition of the input signal, so that the output signal is the inverse of the inhibiting input signal.
It is possible that the output neurons of an inversion nucleus with internal single signal formation take over the role of single signal formers and output neurons at the same time. In this case there is no division into a magnocellular and a parvocellular part.
Where do you find inversion nuclei in the brain? For this purpose, one should be aware of the criteria by which these nuclei can be identified.
Detection criteria for inversion cores with external input signal:
- Supply of excitatory permanent signals (single signals) from a single-signal nucleus to the out-put neurons, usually close to the cell body or the axon mound because of the necessary strong excitation
- Delivery of inhibitory signals, which are temporarily active, to the outer edge of the dendrite tree or to the dendrite branches of the output neurons. Docking the inhibitory axons to the cell body or to the axon mound would not cause relative but total inhibition. In this case a negation nucleus would be present.
- One original neuron of the signal to be inverted supplies one output neuron each.
- A single-signal-providing neuron can indeed supply several (or even many) output neurons simultaneously - the single signal can therefore affect several neurons by splitting it before use. In this case, significantly fewer input signal lines are needed than output lines.
Theoretically inversion cores could also work with negative single signals, their output would then be inhibitory instead of excitatory. However, the author did not find any real example of this.
Detection criteria for inversion cores with internal single signal:
- Supply of excitatory signals, which often pass through the inversion nucleus, being tapped by the output neurons. The neurons that deliver these signals are usually magnocellular mean neurons, e.g. from layer V of the cortex.
- The single-signal-forming neurons are large, magnocellular main neurons, which tap the excitatory input that is introduced or passed through with a huge dendrite tree and form a single signal (permanent signal) from it.
- The single-signal-forming neurons of the inversion nucleus with internal single-signal formation form the magnocellular part of the inversion nucleus.
- Supply of inhibitory input from another region. Each inhibiting axon usually inhibits exactly one output neuron, so that its continuous signal is relatively suppressed during the inhibition phase and therefore inverted.
- Input activity and output activity are inverse to each other, this is true even in small subareas.
- Often the single-signal-forming input is passed through the core and leaves it again relatively unchanged in the direction of its target areas.
Depending on whether the output of an inversion kernel is excitatory or inhibitory, we speak of a positive or negative inversion kernel.
Inversion nuclei and negation nuclei are analogous. In the case of the negation nucleus, however, the inhibitory input has a very strong inhibitory effect, because it acts at the axon mound or directly at the cell body. At the inversion nucleus, the inhibiting synapses are located farther out, where the inhibition is not total, but only relative. Thus the input is not negated but inverted.
While the output neurons of a negation nucleus only know two states (active = one signal, inactive = zero signal), the firing rates of the output neurons of an inversion nucleus assume all values f between a minimum value fmin and a maximum value fmax.
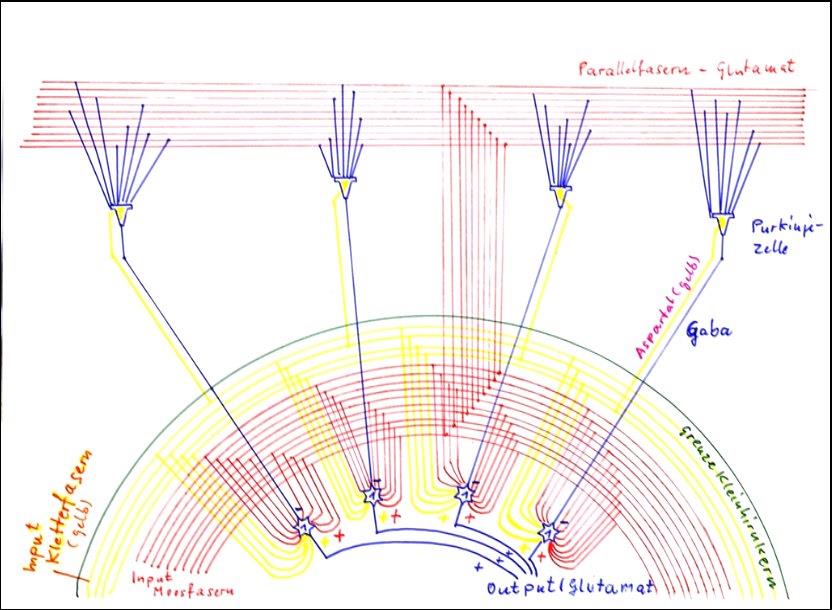
Theorem 1.12: The cerebellar nuclei are positive inversion nuclei for Purkinje cell signals
The cerebellar nuclei are positive inversion nuclei with internal single signal formation. The excitatory signals required for the formation of the single signal originate from the moss fibres on the one hand and the climbing fibres on the other hand. Both types of fibers give off excitatory collaterals in the cerebellar nuclei, which permanently excite the output neurons of the cerebellar nuclei.
The Purkinje cells, on the other hand, produce more or less strong inhibitory signals when excited, the strength of which depends on the current excitation strength of the Purkinje cells. These have a constantly acting, medium inhibitory effect, which is additionally and gradually altered when signals are applied to the parallel fibres or climbing fibres. The author interprets these Purkin signals as inhibitory off-signals. By a relative inhibition of the input signals of the output neurons of the cerebellar nuclei, these signals are inverted at the input signal. Thus the inverted on-signals are created in addition to the off-signals.
Since the output neurons are glutamaterg*, their on signals are excited. Therefore, the cerebellar nuclei are positive inversion nuclei for the negative OFF signals of the Purkinje cells and invert the Purkinje signals. *there are also GABAergic output neurons!
However, the theorem 1.12 only becomes really understandable when the functioning of the cerebellum is explained. It is shown that each Purkinje cell has its own, unmistakable imprinting signal. In particular, it will then be shown that the Purkinje cell responds to an input signal with an inhibitory output whose signal strength decreases the more similar the current parallel fibre signal is to the embossing signal of the Purkinje cell. Therefore, the Purkinje output is always an off-signal whose rate of fire is significant.
Sketch 1.17: The cerebellar nuclei as positive inversion nuclei with internal single signal formation
Monografie von Dr. rer. nat. Andreas Heinrich Malczan