Theory of the brain’s neural circuitry
and analytical thinking
ISBN 978-3-00-037458-6 - ISBN 978-3-00-042153-2
Part 1.2 Typification of neural core regions
For the following explanations we need some elementary basic terms, which will now be defined.
Definition 1.3: Core areas
Accumulations of neurons are called core areas.
Core areas can be distinguished according to the number of input and output lines.
Definition 1.4: Switching cores
The number of input lines is about the same as the number of output lines, but the input and output usually use different transmitters, so that the switchover from one transmitter to another takes place.
Definition 1.5: Convergence nuclei
The number of input lines is significantly greater than the number of output lines.
Definition 1.6: Divergence nuclei or divergence layer
The number of input lines is significantly smaller than the number of output lines.
Under input or output lines we may imagine the axons of the nerve cells that serve to transmit signals. We use the terms switch core, convergence core and divergence core to classify the neural cores.
Theorem 1.2: The substantia nigra pars compacta is a switching core for the cortex output
The substantia nigra pars compacta receives on the one hand the output of the cluster neurons and on the other hand the output of the activity neurons. Both types of output are glutamatergic and are switched to the transmitter dopamine. While the cluster neurons are switched to dopamine of the subtype D2 and inhibit their target neurons in the matrix of the striatum, the activity neurons of the cortex clusters are switched to dopamine of the subtype D1 and excite their target neurons in the striosomes of the striatum.
The substantia nigra pars compacta is a neuronal nucleus of the basal ganglia system. The theorem 1.2 assumes that the main task of the substantia nigra pars compacta is to switch the dopaminergic input from the cortex to the transmitter dopamine. This makes sense because the output of this nucleus is sent back to the cortex, quasi in the direction from which it originally came. However, because it was switched to another transmitter before the direction change, the neural system can easily distinguish between cortex output and nigral output.
However, theorem 1.2 also assumes (an already known) specification of the neurons of the substantia nigra pars compacta in two groups. One group uses the D1 receptors in the target area, the stria-tum, which are excited by the transmitter dopamine. We will refer to this group as dopaminergic neurons of the subtype D1. The other group of neurons of the nuclear region also uses dopaminergic but inhibitory D2 receptors in the striatum. This group of neurons will be referred to as dopaminergic neurons of subtype D2.
The theorem 1.2 assumes a cause for the known compartmentalization of the striatum into the matrix and striosomes. It is known (in 2011) that the dopaminergic axons from the substantia nigra pars compacta have an excitatory effect via D1 receptors and an inhibitory effect via D2 receptors.
The theorem 1.2 assumes a new fact of this compartmentation which has not yet been recognized. It is claimed that the activity neurons of all clusters of the cortex cortex in the substantia nigra pars compacta are switched to dopaminergic neurons of the subtype D1 and subsequently excite the neurons of the striosomes of the striatum, while the signal neurons of all clusters of the cortex in the substantia nigra pars compacta are switched to dopaminergic neurons of the subtype D2 and subsequently inhibit the neurons of the matrix of the striatum.
The deeper meaning of such a complementary projection and the compartmentalization of the striatum into matrix and striosomes is comprehensively elucidated in this theory for the first time.
Sketch 1.2: Substantia nigra as switch core from glutamate to dopamine
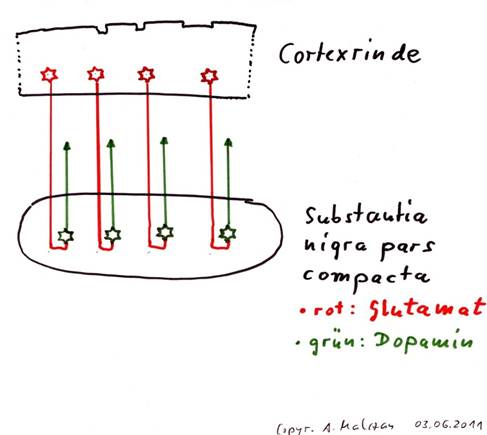
Switching nuclei perform a very important task in the brain, although they only switch a signal from one transmitter to another transmitter. This makes it possible, however, to distinguish between input and output. If the entire nervous system were to use only one type of transmitter, a total signal chaos would result, since each neuron would be transmitter compatible with each neuron. Only the use of different transmitters makes it possible for a neuron to tap only those axons from a huge range of signals to which it is transmitter-compatible.
Theorem 1.3: The striosome system of the striatum is a switch core
The activity neurons (mean neurons) of the cortex clusters project excitedly into the switch core substantia nigra pars compacta, where they are switched to dopamine of subtype D1. This output is switched to the transmitter GABA in the striosomes of the striatum. Therefore, the striosome system is a switching core.
It remains to be seen whether the matrix system of the striatum has other functions.
Theorem 1.4: The nucleus subthalamicus is a neuronal convergence nucleus
The subthalamic nucleus receives the output of the activity neurons of the cortex clusters. From the average signal levels delivered, its neurons form area averages by a renewed averaging process, which represent the activity state of larger cortex regions, which themselves consist of many cortex clusters.
In this respect, the output neurons of the nucleus subthalamicus are second stage activity neurons. Their output is usually a more or less strong continuous signal whose rate of fire represents the area activity of the corresponding cortex area.
It should also be noted that the cortex cortex often receives complementary signals, i.e. those in which there is both an on-variant and an off-variant (e.g. in the visual primary cortex). Thus, the activity neuron of the subthalamic nucleus almost always receives an input. Only if large cortex areas are completely without signal, the output neurons of the subthalamicus could also remain silent.
Sketch 1.3: The nucleus subthalamicus as convergence nucleus for the cortex output from layer V
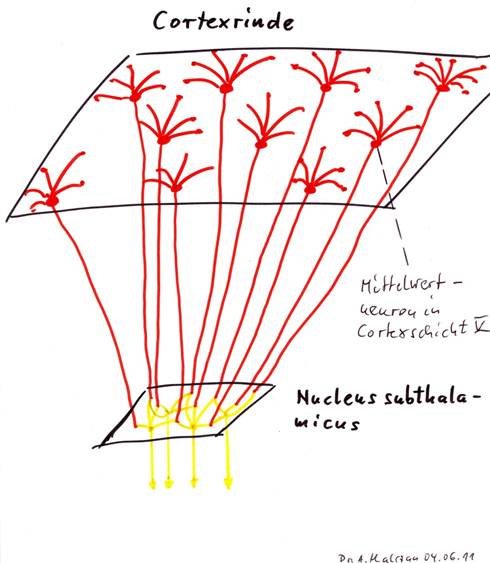
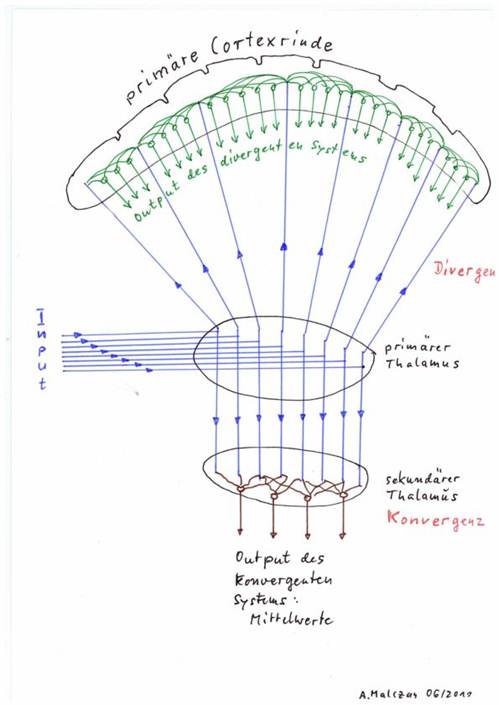
Convergence cores and divergence cores can of course also be
switchover cores. It must be pointed out that ultimately the receptors of the
ion channels decide whether a transmitter has an excitatory or inhibitory
effect. For many transmitters there are several types of receptors with
different effects.
This theory also aims to show that the brain works like a computer. Therefore it is necessary to introduce some elementary basic concepts from the computer world.
Definition 1.7: Zero signal
We assign the rate of fire f = 0 to a neuron that does not produce action potentials and call its output a zero signal. It then has the signal value zero.
Definition 1.8: Single signal
We assign the input signal to a neuron that generates action potentials with an arbitrary rate of fire f greater than zero. It has the signal value 1.
Thus an actually firing neuron has the signal value 1, while a non firing neuron has the signal value zero. Thus, neurons represent a binary signal value according to the "all or nothing rule". This is the first indication of computer-like behavior.
Sketch 1.5: Neural single signal - not necessarily frequency stable:

Neurologists use to refer to accumulations of nerve cells as nuclei. The nucleus
subthalamicus is such a nucleus. Its output axons (usually) carry a continuous
signal, which is produced by double averaging over the cluster quantity of the
cortex. However, since continuous signals are generated by actually firing
neurons and their (binary) signal value is 1, the output neurons of the nucleus
subthalamicus ultimately generate single signals. However, these signals usually
have different firing rates.
Definition 1.9: Positive single-signal core
If a neuron nucleus generates excitatory continuous signals (single signals) from the excitatory mean value signals supplied to it or passing through it, it is called a positive single-signal nucleus.
Definition 1.10: Negative single-signal core
If a neuron nucleus generates inhibitory continuous signals (single signals) from the excitatory mean value signals supplied to it or passing through it, it is called a negative single-signal nucleus.
Theorem 1.5: The nucleus subthalamicus is a positive single-signal nucleus
The nucleus subthalamicus is a positive single signal nucleus. In most cases, its output neurons produce excitatory single signals.
Sketch 1.6: Origin of single signals (continuous signals) from an uninterrupted sequence of action potentials

Changed time resolution:
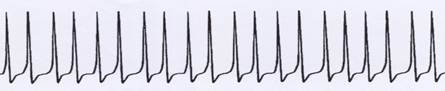
Even more time-stamped:
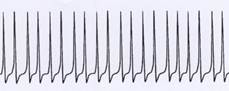
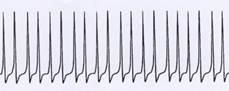
Very tightly packed in time: Example of a single signal (continuous signal)





However, if in special cases (sleep, deep fainting, coma) the cortex activity is strongly reduced in certain larger regions, part of the output of the subthalamic nucleus may also be the zero signal.
Neurologists may be surprised that a neuronal continuous signal is called a "one signal", whereas a non-existent signal is called a "zero signal". But a neural number theory also requires the basic elements of number theory. The zero element and the one element are the basis of all number spaces.
Furthermore, zero and one as numerical equivalents of the logical values "True" and "False" are important basic elements of propositional logic and enable binary calculation.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan