Vertebrate brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-064888-5
7.4 Color analysis in plane divergence grids
In this section, we investigate the consequences of a plane divergence grating receiving visual color signals. Here, the signals are distributed in a plane by divergence. The development of the color sense can be explained particularly well with a plane divergence grating. It remains to be seen whether these flat divergence grids are located in the visual cortex, in the amygdala or in the nucleus olivaris. We hypothetically assume that the color analysis is performed in the nucleus olivaris. If we were to choose the visual cortex or the amygdala, there would be analog signaling pathways that also exist in reality.
Parallel to the brightness analysis, a very fine color analysis was also possible. Coming from the retina, partly running directly via the tectum, partly indirectly via the corpus geniculatum laterale to the tectum, the colour signals reach the nucleus olivaris via the tractus tectoolivaris and from there as climbing fibres the cerebellum.
Some known facts can be used, but some assumptions need to be confirmed by further research.
With regard to color vision, we examine here as an example the case of primates, where the colors red, green and blue can be evaluated. In this example the retinal ganglion cells provide the red signal red-on/green-off, furthermore the green signal green-on/red-off and a blue signal blue-on/yellow-off.
We theoretically divide the retina into pixels and have these three color signals for each pixel. This is a simplification, because the density of ganglion cells is not constant within the retina.
Each retinal pixel is now assigned three associated ganglion cells, one for the red signal, one for the green signal and one for the blue signal. We imagine these ganglion cells arranged in the retinal plane in such a way that they form a triangle that belongs to the image pixel. We call this triangle of these ganglion cells the retinal color triangle of the pixel.
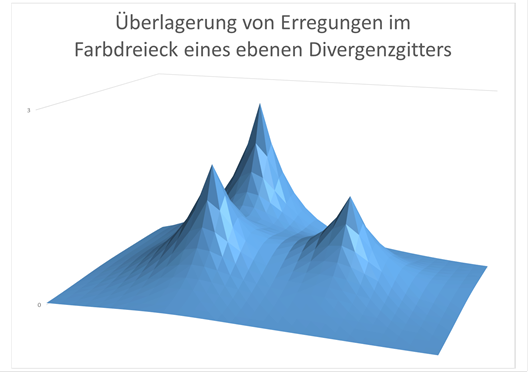
Figure 75 - Superposition of the excitations in a color triangle
Figure:
Schematic representation of the excitation generated by superposition in a color triangle of the nucleus olivaris, the retinal excitation arrives where the excitation maxima are located. The excitations of the neighbouring colour triangles have been omitted for simplification. The three different maxima represent the three relative fire rates of the corresponding color neurons in the olivar nucleus. In white, the three "peaks" are equal in height.
The signaling pathway of the color signals to the cerebellum should be via the nucleus olivaris as described. This nucleus was already in early evolutionary history an interference nucleus for the signals of different modalities and served mainly for motor body control depending on the input of the different modalities. We therefore assume that the visual signals reached it just as much as the vestibular or motor signals. Initially, there was certainly only one type of color receptor in the retina. Their signals went to the nucleus olivaris. When new colour receptors were formed through mutation, their axons took the same path to the olive nucleus.
The color signal output of the retina should therefore reach the nucleus olivaris in a topologically well-ordered manner. Each color ganglion cell of the retina projects via the tectum into an assigned color neuron of the nucleus olivaris. The retinal red signal thus reaches a red neuron, the green signal a green neuron and the blue signal a blue neuron. Thus, a color triangle is formed in the nucleus olivaris for each retinal pixel depicted, the corners of which are formed by the corresponding color neurons. Then this area of the nucleus olivaris looks like a section of an LCD screen, which uses red, green and blue colour diodes.
Initially (but only after the formation of the different color receptors) the color sensitive part of the nucleus olivaris was thus approximately a 1:1 copy of the retinal color sensitive ganglion cells of the existing color classes. Three projection neurons for each retinal pixel form a colour triangle red-green-blue and guide the retinal colour output as climbing fibres to the cerebellum.
However, as with spinal input, the process of redundancy generation through signal divergence occurred very early in the history of evolution. In the nucleus olivaris, additional projection neurons were formed (for each image pixel) between the original three projection neurons of the primary colors red, green, and blue in order to achieve redundancy. This allowed individual projection neurons to die without significantly impeding signal evaluation. This was associated with an increase in area, because now there were no longer three neurons per image pixel, which formed a triangle, but 6, 10, 15, etc. If one side of the triangle consisted of n neurons, the extended color triangle now consisted of (n+1)*n/2 neurons. The input reached each of these now much larger color triangles only at the corners of the triangle, the number of input neurons remained the same. But the number of output neurons grew excessively. To simplify, let's imagine one output neuron in each triangle corner where the input also arrives.
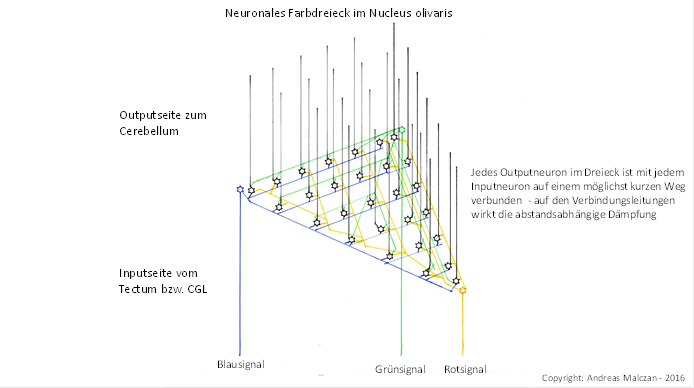
Figure 76 - Neural color triangle in the olivar nucleus
And as already described, a distance-dependent attenuation occurred in the nucleus olivaris. The further an output neuron of the extended color triangle was away from one of the three input neurons of its image pixel, the stronger the distance-dependent exponential attenuation of the corresponding color signal became. This is because the nucleus olivaris formed a grey nucleus in which the axons and dendrites were unmyelinated. The excitation propagation was subject to the cable equation for non-myelinated fibers.
If we think of the fire rate of the three color components in the corners of the associated triangle plotted as a function value of a two-dimensional function, there is a kind of mountain peak in each input neuron that we could think of in the associated color. The excitation decreased exponentially with increasing distance from this peak. Between the three maximum values in the corners of the color triangle of an image pixel there was usually a minimum value whose position depended on the fire rate ratio of the three color signals involved. In a model, there was a valley (the minimum) between the red, green and blue mountain tops, and the higher one of these color mountains was, the further away from it the valley was. Thus, the minimum of neuronal excitation in the extended color triangle of the nucleus olivaris encoded the red-green-blue color ratio and thus the corresponding mixed color of the additive color mixture for the respective pixel of the retina. If all three colour signals were of equal strength, i.e. had the same fire rates, the minimum was in the centre of the triangle and was assigned to the colour white or grey. Each output neuron of the extended color triangle was thus assigned to a color from the normal color spectrum (of the rainbow). And the projection neuron with the minimum excitation encoded in each color triangle of the nucleus olivaris the real color of light acting on the retinal pixel.
However, there was an excitation minimum even if one or even two primary colours red, green or blue were completely absent. And if it was the representation of a colored area, all corresponding color triangles, which were represented on the nucleus olivaris, were inside this area in such a way that there was a signal minimum for each color triangle as long as any color signal was present. If only a single color triangle had been driven with the signal of a base color, while the neighboring color triangles received no input, the minimum could also be located outside this triangle area. But if the neighboring triangles also received input, a minimum outside the triangle in question became impossible because it was prevented by the input there.
Only at the outline of a color area the described color evaluation could not be effective via minimum coding.
However, the minimum coding was not so easy to use for the vertebrate. If a certain colour or mixture of colours were to trigger a motor response, i.e. actively control muscles, maximum excitation would be more favourable than minimum excitation. Therefore, the minimum coded colour signals of the nucleus olivaris had to be converted into maximum coded signals by signal inversion.
However, signal inversion was - at least in the early evolutionary history of vertebrates - a specialty of spinocerebellum. Traditionally, the spinocerebellum received the output of the nucleus olivaris as input. The excitations of the projection neurons of each color triangle of the nucleus olivaris were topologically sorted and fed to the cerebellum via climbing fibers and inverted there. The mean value signal necessary for the signal inversion was derived from the magnocellular output of the light-on-ganglion cells, which were already mean values of the red, green and blue color signals. And since the receptor density was at its highest in the centre of the retina, the yellow spot, the assigned mean area was relatively small, while it continued to increase radially outwards. The signal used for averaging thus originated from an average area of the retina whose radius was smallest in the yellow spot and became larger radially outwards. This had consequences for the imaging errors of the system.
The output of the cerebellum was a maximum coded signal for each of the color triangles, which (among other things) reached the thalamus in the higher vertebrates. However, this thalamus was not the Corpus geniculatum laterale, as this received its input directly from the retina without the cerebellum being interposed. We want to call this new thalamus region the secondary visual thalamus. It received the visual cerebellum output.
The thalamus with its many GABAergic interneurons provided strong lateral inhibition within the signals of each color triangle and caused (ideally) only one thalamic output neuron in each triangle to be active and represent the signal strength maximum, while the remaining output neurons were (almost) completely inhibited.
Thus the color of the light falling on the retinal pixel had been digitized, because only one visual thalamic neuron per pixel was active, and for each color mixture it was different. For each retinal image pixel there was an axon bundle in which the signals of the corresponding color triangle encoded the color in maximum code. And the finer the color resolution, the more axons belonged to such an axon bundle.
The output of the secondary thalamus moved to the frontal lobe and reached its own area where the digitized color signals ended. Whether the topology of the color triangles was preserved or a new sorting, for example according to retinal coordinates and colors, may be clarified by research. In any case, this area must have significantly more neurons than the retina. The finer the cerebellar colour resolution is, the more neurons this cortical colour evaluation area must have.
The output of this area in the frontal lobe reached other projection areas. On the one hand, intracortical projection fibers reached the other lobi, especially the higher visual areas MT and V4, but there was also the strong projection of this cortex area back to the cerebellum. The output of this cortex area is supplied to the pontocerebellum via the bridge cores and enables the learning of color objects.
Furthermore, this output reaches the limbic system and is buffered there by the signal of each active color neuron of the secondary thalamus participating in the limbic rotation in each color triangle. From there, this rotating signal (according to the author's theory) again reaches the nucleus olivaris and docks exactly to the original neuron. As long as the signal rotation continues in the corresponding limbic signal loop, the corresponding neuron in the nucleus olivaris receives its excitation and therefore also fires.
It has not yet been proven that there are indeed plane divergence grids in the nucleus olivaris. Some reasons speak against it, others for it. Visual input into the cerebellum has been proven. But it is not difficult to shift the presented model of plane divergence grids for optical color signals from the nucleus olivaris to the visual cortex cortex. The cortex cortex of the primary regions is certainly organized in plane divergence grids. Therefore, the presented model may also have been realized in the visual cortex cortex. This is also true for the phenomena of color afterimages, the color triangle and color constancy presented in the following chapters. It is therefore possible that all these phenomena do not take place in the nucleus olivaris, but in the visual cortex cortex cortex. The algorithm involved remains the same. Nevertheless, in the author's opinion, color processing is mainly realized in the cerebellum.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan