Gehirntheorie der Wirbeltiere
ISBN
978-3-00-064888-5
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
7.4 Die Farbanalyse in ebenen Divergenzgittern
In diesem Abschnitt untersuchen wir, welche Folgen es hat, wenn ein ebenes Divergenzgitter visuelle Farbsignale empfängt. Hier werden die Signale durch Divergenz in einer Ebene verteilt. Die Entwicklung des Farbsinnes ist mit einem ebenen Divergenzgitter besonders gut erklärbar. Es sei an dieser Stelle dahingestellt, ob sich diese ebenen Divergenzgitter im visuellen Cortex, in der Amygdala oder im Nucleus olivaris befinden. Wir nehmen hypothetisch an, die Farbanalyse erfolge im Nucleus olivaris. Würden wir uns für den visuellen Cortex oder die Amygdala entscheiden, gäbe es analoge Signalwege, die ebenfalls real existieren.
Parallel zur Helligkeitsanalyse wurde auch eine sehr feine Farbanalyse möglich. Von der Retina kommend, teils direkt über das Tectum verlaufend, teils indirekt über das Corpus geniculatum laterale zum Tectum ziehend, erreichen die Farbsignale über den Tractus tectoolivaris den Nucleus olivaris und von dort als Kletterfasern das Cerebellum.
Hierbei können einige bekannte Fakten verwendet werden, einige Annahmen sind jedoch durch weitere Forschung zu bestätigen.
Bezüglich des Farbsehens untersuchen wir hier beispielhaft den Fall der Primaten, bei denen die Farben Rot, Grün und Blau ausgewertet werden können. Die Ganglienzellen der Netzhaut liefern in diesem Beispiel das Rotsignal Rot-On/Grün-Off, weiterhin das Grünsignal Grün-On/Rot-Off und ein Blausignal Blau-On/Gelb-Off.
Wir unterteilen die Netzhaut theoretisch in Pixel und haben für jedes Pixel jeweils diese drei Farbsignale. Dies ist eine Vereinfachung, denn die Dichte der Ganglienzellen ist innerhalb der Netzhaut nicht konstant.
Jedem Netzhautpixel sind nun drei zugehörige Ganglienzellen zugeordnet, eine für das Rotsignal, eine für das Grünsignal und eine für das Blausignal. Wir denken uns modellhaft diese Ganglienzellen in der Netzhautebene so angeordnet, dass sie ein Dreieck bilden, welches zum Bildpixel gehört. Dieses Dreieck aus diesen Ganglienzellen bezeichnen wir als das retinale Farbdreieck des Pixels.
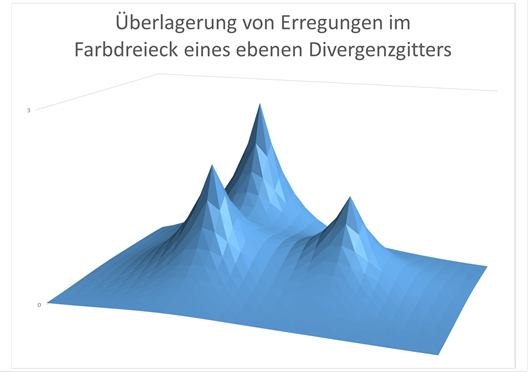
Abbildung 75- Überlagerung der Erregungen in einem Farbdreieck
Abbildung:
Schematische Darstellung der durch Überlagerung entstehenden Erregung in einem Farbdreieck des Nucleus olivaris, die retinale Erregung trifft dort ein, wo die Erregungsmaxima liegen. Die Erregungen der benachbarten Farbdreiecke sind vereinfachend weggelassen worden. Die drei unterschiedlichen Maxima stellen die drei relativen Feuerraten der zugehörigen Farbneuronen in Nucleus olivaris dar. Bei Weiß sind die drei -Bergspitzen- gleich hoch.
Der Signalweg der Farbsignale zum Cerebellum möge wie beschrieben über den Nucleus olivaris erfolgen. Dieser Kern war bereits in früher Evolutionsgeschichte ein Interferenzkern für die Signale verschiedener Modalitäten und diente vornehmlich der motorischen Körpersteuerung in Abhängigkeit vom Input der verschiedenen Modalitäten. Wir unterstellen also, die visuellen Signale hätten ihn ebenso erreicht wie die vestibulären oder die motorischen Signale. Anfangs war sicherlich nur eine Art von Farbrezeptoren in der Netzhaut vorhanden. Deren Signale zogen zum Nucleus olivaris. Als sich durch Mutation neue Farbrezeptoren bildeten, nahmen deren Axone den gleichen Weg zum Olivenkern.
Der Farbsignaloutput der Netzhaut möge also den Nucleus olivaris topologisch wohlgeordnet erreichen. Jede Farbganglienzelle der Netzhaut projiziere über das Tectum in ein zugeordnetes Farbneuron des Nucleus olivaris. Das retinale Rotsignal erreicht also ein Rot-Neuron, das Grünsignal ein Grün-Neuron und das Blau-Signal ein Blau-Neuron. So entsteht auch im Nucleus olivaris zu jedem abgebildeten Netzhautpixel ein Farbdreieck, dessen Ecken von den zugehörigen Farbneuronen gebildet werden. Dann sieht diese Fläche des Nucleus olivaris aus wie ein Ausschnitt aus einem LCD-Bildschirm, der rote, grüne und blaue Farbdioden verwendet.
Anfangs (jedoch erst nach Herausbildung der verschiedenen Farbrezeptoren) war der farbsensitive Teil des Nucleus olivaris also in etwa eine 1:1-Kopie der retinalen farbempfindlichen Ganglienzellen der vorhandenen Farbklassen. Je drei Projektionsneuronen für jedes Netzhautpixel bilden ein Farbdreieck Rot-Grün-Blau und leiten den retinalen Farboutput als Kletterfasern zum Cerebellum.
Doch wie bereits beim spinalen Input erfolgte auch hier recht früh in der Evolutionsgeschichte der Prozess der Redundanzerzeugung durch Signaldivergenz. Im Nucleus olivaris bildeten sich (für jedes Bildpixel) zwischen den ursprünglich drei Projektionsneuronen der Primärfarben Rot, Grün und Blau zusätzliche Projektionsneuronen, um eine Ausfallsicherheit zu erzielen. So konnten einzelne Projektionsneuronen absterben, ohne dass die Signalauswertung wesentlich behindert wurde. Verbunden war dies mit einer Flächenzunahme, denn nun gab es pro Bildpixel nicht mehr drei Neuronen, die ein Dreieck bildeten, sondern 6, 10, 15, usw. Bestand eine Dreieckseite aus n Neuronen, so bestand das erweiterte Farbdreieck nunmehr aus (n+1)*n/2 Neuronen. Der Input erreichte jedes dieser nun wesentlich größeren Farbdreiecke nur an den Ecken des Dreiecks, die Anzahl der Inputneuronen blieb ja gleich groß. Aber die Anzahl der Outputneuronen wuchs exzessiv. Vereinfach denken wir uns auch je ein Outputneuron genau in jeder Dreiecksecke, wo auch der Input eintrifft.
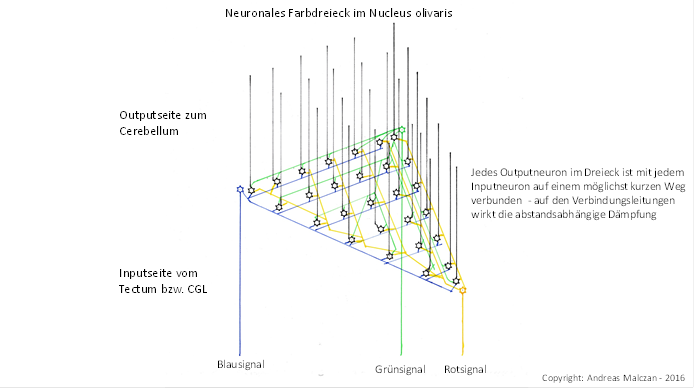
Abbildung 76 - Neuronales Farbdreieck im Nucleus olivaris
Und wie bereits beschrieben, trat im Nucleus olivaris eine abstandsabhängige Dämpfung auf. Je weiter ein Outputneuron des erweiterten Farbdreiecks von einem der drei Inputneuronen seines Bildpixels entfernt war, umso stärker wurde die abstandsabhängige exponentielle Dämpfung des entsprechenden Farbsignals. Denn der Nucleus olivaris bildete ja einen grauen Kern, in dem die Axone und Dendriten unmyelinisiert waren. Die Erregungsausbreitung unterlag der Kabelgleichung für marklose Fasern.
Denken wir uns die Feuerrate der drei Farbanteile in den Ecken des zugehörigen Dreiecks als Funktionswert einer zweidimensionalen Funktion aufgetragen, so gibt es in jedem Inputneuron eine Art Bergspitze, die wir uns in der zugehörigen Farbe denken könnten. Die Erregung nahm mit zunehmendem Abstand von dieser Spitze exponentiell ab. Zwischen den drei Maximalwerten in den Ecken des Farbdreiecks eines Bildpixels lag meist ein Minimumwert, dessen Lage vom Feuerratenverhältnis der drei beteiligten Farbsignale abhing. Modellhaft lag zwischen der roten, der grünen und der blauen Bergspitze ein Tal (das Minimum), und je höher einer dieser Farbberge war, umso weiter war das Tal von ihm entfernt. Somit codierte das Minimum der neuronalen Erregung im erweiterten Farbdreieck des Nucleus olivaris das Farbverhältnis Rot-Grün-Blau und damit die zugehörige Mischfarbe der additiven Farbmischung für den betreffenden Bildpunkt der Retina. Waren alle drei Farbsignale gleichstark, hatten also die gleichen Feuerraten, so lag das Minimum in der Dreiecksmitte und war der Farbe Weiß bzw. Grau zugeordnet. Jedes Outputneuron des erweiterten Farbdreiecks war somit einer Farbe aus dem normalen Farbspektrum (des Regenbogens) zugeordnet. Und das Projektionsneuron mit der minimalsten Erregung verschlüsselte in jedem Farbdreieck des Nucleus olivaris die real auf das Netzhautpixel einwirkende Lichtfarbe.
Ein Erregungsminimum gab es jedoch auch bei völligem Fehlen einer oder gar zweier Primärfarben Rot, Grün bzw. Blau. Und wenn es sich um die Abbildung einer farbigen Fläche handelte, so waren alle zugehörigen Farbdreiecke, die auf dem Nucleus olivaris abgebildet wurden, im Inneren dieser Fläche so beschaffen, dass zu jedem Farbdreieck ein Signalminimum gab, solange irgendein Farbsignal vorhanden war. Wäre nur ein einzelnes Farbdreieck mit dem Signal einer Grundfarbe angesteuert worden, während die benachbarten Farbdreiecke keinen Input erhielten, so könnte das Minimum auch außerhalb dieser Dreiecksfläche liegen. Wenn aber die Nachbardreiecke ebenfalls Input bekamen, wurde ein Minimum außerhalb des betreffenden Dreiecks unmöglich, weil es vom dortigen Input verhindert wurde.
Nur am Umriss einer Farbfläche konnte die beschriebene Farbauswertung über eine Minimumcodierung nicht wirksam werden.
Die Minimumcodierung war jedoch für das Wirbeltier nicht so gut zu verwerten. Sollte eine bestimmte Farbe oder Farbmischung eine motorische Reaktion auslösen, also Muskeln aktiv ansteuern, so wäre eine maximale Erregung günstiger als eine minimale. Daher mussten die minimumcodierten Farbsignale des Nucleus olivaris durch Signalinversion in maximumcodierte Signale umgewandelt werden.
Die Signalinvertierung war jedoch - wenigstens in früher Evolutionsgeschichte der Wirbeltiere - eine Spezialität des Spinocerebellums. Dieses erhielt ja traditionell den Output des Nucleus olivaris als Input zugeführt. Die Erregungen der Projektionsneuronen eines jeden Farbdreiecks des Nucleus olivaris wurden topologisch sortiert dem Cerebellum über Kletterfasern zugeleitet und dort invertiert. Das für die Signalinversion nötige Mittelwertsignal wurde aus dem magnocellularen Output der Hell-On-Ganglienzellen abgeleitet, die ja bereits Mittelwerte der roten, grünen und blauen Farbsignale waren. Und da die Rezeptorendichte in Zentrum der Netzhaut, dem gelben Fleck, an größten war, war das zugeordnete Mittelwertgebiet relativ klein, während es weiter radial nach außen größer wurde. Das zur Mittelwertbildung verwendete Signal entstammte also einer Mittelwertfläche der Netzhaut, deren Radius im gelben Fleck am kleinsten war und nach außen größer wurde. Das hatte Folgen für die Abbildungsfehler des Systems.
Der Output des Cerebellums war für jedes der Farbdreiecke ein maximumcodiertes Signal, welches (unter anderem) bei den höheren Wirbeltieren den Thalamus erreichte. Dieser Thalamus war jedoch nicht das Corpus geniculatum laterale, denn dieses erhielt seinen Input direkt von der Retina ohne Zwischenschaltung des Cerebellums. Wir wollen dieses neue Thalamusgebiet als sekundären visuellen Thalamus bezeichnen. Er erhielt den visuellen Cerebellumoutput.
Der Thalamus mit seinen vielen GABAergen Interneuronen sorgte für eine starke laterale Hemmung innerhalb der Signale eines jeden Farbdreiecks und bewirkte, dass (idealerweise) nur noch ein thalamisches Outputneuron in jedem Dreieck aktiv war und das Signalstärkemaximum repräsentierte, während die übrigen Outputneuronen (fast) völlig gehemmt wurden.
Somit war die Farbe des auf das Netzhautpixel fallenden Lichtes digitalisiert worden, denn nur noch ein visuelles Thalamusneuron pro Pixel war aktiv, und für jede Farbmischung war es ein anderes. Zu jedem Bildpixel der Netzhaut gab es ein Axonbündel, in dem die Signale des zugehörigen Farbdreiecks maximumcodiert die Farbe verschlüsselten. Und je feiner die Farbauflösung war, umso mehr Axone gehörten zu einem solchen Axonbündel.
Der Output des sekundären Thalamus zog zum Frontallappen und erreichte dort ein eigenes Gebiet, in dem die digitalisierten Farbsignale endeten. Ob dabei die Topologie der Farbdreiecke erhalten blieb oder eine Neusortierung etwa nach Retinakoordinaten und Farben erfolgte, möge die Forschung klären. Auf alle Fälle muss dieses Gebiet im Vergleich zur Retina deutlich mehr Neuronen aufweisen. Je feiner die cerebellare Farbauflösung ist, umso mehr Neuronen muss dieses cortikale Farbauswertungsgebiet besitzen.
Der Output dieses Gebietes im Frontallappen erreichte weitere Projektionsgebiete. Einerseits wurden über intracortikale Projektionsfasern die anderen Lobi erreicht, vor allem die höheren visuellen Areale MT und V4. Aber es gab auch die starke Projektion dieses Cortexgebietes zurück zum Cerebellum. Der Output dieses Cortexgebietes wird über die Brückenkerne dem Pontocerebellum zugeführt und ermöglicht das Erlernen von Farbobjekten.
Weiterhin erreicht dieser Output das limbische System und wird dort zwischengespeichert, indem das Signal eines jeden aktiven Farbneurons des sekundären Thalamus in jedem Farbdreieck an der limbischen Rotation teilnimmt. Von dort erreicht dieses rotierende Signal (nach der Theorie des Autors) wieder den Nucleus olivaris und dockt genau am Ursprungsneuron an. Solange die Signalrotation in der zugehörigen limbischen Signalschleife andauert, erhält das zugehörige Neuron im Nucleus olivaris dessen Erregung zugeführt und feuert daher ebenfalls.
Es ist gegenwärtig noch nicht nachgewiesen, dass es im Nucleus olivaris tatsächlich ebene Divergenzgitter gibt. Einige Gründe sprechen dagegen, andere dafür. Visueller Input ins Cerebellum ist nachgewiesen worden. Aber es bereitet keinerlei Schwierigkeiten, das vorgestellte Modell der ebenen Divergenzgitter für optische Farbsignale vom Nucleus olivaris zur visuellen Cortexrinde zu verlegen. Die Cortexrinde der primären Gebiete ist mit Sicherheit in ebenen Divergenzgittern organisiert. Daher kann das vorgestellte Modell auch in der visuellen Cortexrinde realisiert worden sein. Dies gilt auch für die nachfolgend vorgestellten Phänomene der farbigen Nachbilder, das Farbdreieck und die Farbkonstanz, die in den folgenden Kapiteln beschrieben werden. Es ist also möglich, dass alle diese Phänomene nicht im Nucleus olivaris, sondern in der visuellen Cortexrinde stattfinden. Der beteiligte Algorithmus bleibt der gleiche. Dennoch wird die Farbverarbeitung nach Ansicht des Autors vorwiegend im Cerebellum realisiert.
Hinweis des Autors nach Drucklegung:
- ← 7.3 Die Helligkeitsanalyse im Cerebellum
- 7.5 Der Farbbegriff und das neuronale Farbdreieck des Nucleus olivaris →
Monografie von Dr. rer. nat. Andreas Heinrich Malczan