Vertebrate brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-064888-5
6. The further development of the striatum and the basal ganglia system as a result of signal divergence in the cortex
6.1 The use of inhibitory inversion neurons for motion detection
The vestibulo- and spinocerebellum used the signal inversion on permanently excited neurons as their main working principle. The signal to be inverted reached the Purkinje cell via the climbing fibre, which switched it to inhibitory transmitters. This signal inhibited a permanently excited neuron in the cerebellar nucleus. The permanent excitation of this neuron originated from the reticular format. The inhibition was relative and increased with the rate of fire. This was exactly the signal inversion. Because the permanently excited neuron in the cerebellar nucleus used the transmitter glutamate, the inverted signal was excited. The output of the cerebellar nucleus was the inverted output of the nucleus ruber and was used for motor control of the opposite side.
A significant development in the nervous system of future vertebrates occurred when permanently excited neurons were used for signal inversion using an inhibitory transmitter. This occurred in GABAergic neurons. Such inversion neurons are found, for example, in the matrix of the striatum, in the globus pallidus, but also in the amygdala.
According to the author, the inhibitory inversion neurons found at many locations in the vertebrate brain have a common genesis, which is summarized in the following theorem.
Theorem of the formation of inhibitory inversion neurons
Inhibitory inversion neurons are used for the signal inversion of signals.
They develop as descendants of inhibitory interneurons, which originally served the purpose of lateral inhibition, i.e. contrast enhancement.
They receive continuous excitation, either from excitatory axons passing by and contacting them (e.g. striatum), or they receive continuous excitation from an average nucleus (e.g. globus pallidus).
You continue to receive an inhibiting input signal. This only inhibits them relatively. As the signal strength of this signal increases, the inhibiting effect increases.
The relative inhibition of the existing permanent excitation led to a signal inversion of the input signal.
It is worth remembering here the theorem of motion detection by the primordial triatum. Dopaminergic neurons received the cortical output of classes 5 and 6 of the first floor of the rope ladder nervous system and excited the striosomal neurons in the striatum. These neurons projected inhibitively into the nucleus ruber. This early tractus tegmentalis centralis, even if it consisted of only a few neurons, already enabled movement recognition and the perception of changes. The excitatory present signals and the inhibitory, time-delayed past signals created a differential mapping that was time-selective.
In the cortex, signal divergence of signal class 3 had established itself in the meantime - after a long development path. Each cortical input neuron of class 4 projected onto a huge number of class 3 neurons, resulting in an enormous expansion of the cortex surface, so that due to lack of space, wrinkling even occurred, which could be used to additionally increase the surface area.
These new signals, generated by strong signal divergence, had to be taken into account in the mean value systems in the same way as the previous ones. The axons of class 3 neurons were ultimately treated in the same way as those of classes 5 and 6, and also projected into the substantia nigra pars compacta. There, each axon was switched back to dopamine while maintaining the rate of fire and sent back towards the striatum to contact a GABAergic striate neuron.
In the striatum itself, however, there was a further development. The GABAergic striosome neurons used the dopamine receptor D2. In the striatum of today's vertebrates there is also the dopamine receptor D1. There must therefore have been such receptors in striatum neurons in early primeval times. We therefore assume the development of striate neurons that possessed the dopamine receptor D1. Neurons with the dopamine receptor D1 are inhibited by dopamine. The new class of striate neurons with the dopamine receptor D1 formed a new structure called matrix.
The matrix neurons developed into inhibitory inversion neurons. They formed large dendrite trees with which they integrated the excitation of the class 5 cortexaxons passing through their territory to form a tonic permanent excitation.
Theorem of tonic excitation of matrix neurons
The matrix neurons were forced to be permanently excited by the Class 5 cortex signals passing through them.
For each matrix neuron there was exactly one assigned class 3 cortex neuron, which excitedly projected into the substantia nigra pars compacta. There the cortex signal was switched to dopamine and sent back to the assigned matrix neuron to inhibit it. This corresponded to the inversion of the cortex signal.
Theorem of the matrix of striatum as inversion core
The matrix of the striatum is an inversion core, which obtains the mean value signal required for inversion from the Class 5 cortex output passing through it. The cortex output of the class 3 neurons is inverted in a point-to-point mapping after it has been switched to dopamine on the detour to the substantia nigra pars compacta, which caused a time delay.
Theorem of the matrix output of early striatum
The output of the matrix of the early striatum of one hemisphere was the inverted, time-delayed and GABA-switched cortex output of the third layer of the same hemisphere.
After this expansion, the striatum represented a double body model of the cortex. The striosomes were a time-delayed mean model of class 6 and a time-delayed signal model of class 5, while the matrix represented an inverse and time-delayed signal model of class 3 neurons. All three models used the inhibitory transmitter GABA.
The original task of the striatum, to form a time-sensitive differential image for motion analysis, could not be achieved with the matrix signals in this way. Although the cortex output was switched to GABA, it had once been inverted.
However, the signals of the class 3 neurons were extreme value coded. The reason for this was the nonlinear signal attenuation on non-markless axons, which distributed the input excitation to the many class 3 output neurons.
If these signals were minimum coded in the cortex, they were maximum coded in the striatum. However, if they were maximum coded in the cortex, they were present in the striatum in minimum coded form. Whether they were minimum coded or maximum coded in the cortex, they had the opposite coding in the striatum. Only a second inversion was able to generate signals suitable for difference mapping. However, the solution was already available.
The axons of the matrix neurons, as well as the axons of the class 5 and 6 cortex neurons, moved downwards towards the nucleus ruber. On their way, they passed the mean nucleus of the thalamic level, which was formed by the nucleus subthalamicus. Its neurons were an average model of the cortical floor and were tonically excited (if sufficient cortical signals were present).
Within the nucleus subthalamicus there were also inhibiting interneurons. They received the input of this nucleus and caused a lateral inhibition, i.e. contrast enhancement in this mean nucleus.
We postulate that the GABAergic matrix neurons were compatible with these inhibitory interneurons, they used the same transmitter. That's why they made synaptic contact. The matrix neurons inhibited these interneurons. However, they also received an exciting influx from the glutamatergic neurons of the subthalamic nucleus. The stronger the gabaerge coupling to the matrix axons, the stronger the coupling to the glutamatergic axons, which were tonic mean excitation. These inhibitory neurons formed an independent nucleus called globus pallidus. Its projection axons sought the path from which their excitation came and moved upwards, where they joined the thalamic floor and contacted neurons there. Thus, the axons of the globus pallidus did not end in the nucleus ruber; their number was obviously too large for this. They moved to the signal-related thalamus. The thalamus in the rope ladder system consisted of the sensory and the motor centre. The axons of the globus pallidus moved to the motor side, where all cortexaxons of the class 3 neurons arrived. They merged with these in a point-to-point mapping and formed the time-sensitive differential mapping.
Theorem of the origin of the Globus pallidus
The globus pallidus is a descendant of the
inhibitory
interneurons of the subthalamic nucleus, which
were responsible for lateral inhibition, i.e. contrast enhancement. They receive
the tonic permanent excitation from this nucleus and are inhibited by the matrix
neurons of the striatum, so that they invert the matrix signals. They project
into thalamic nuclei, into which they deliver the inhibitory component to a
time-sensitive differential image.
The following algorithm had thus been established:
- cortex neuron projected into a neuron of the substantia nigra pars compacta.
- This dopaminergic neuron switched the cortical signal to the transmitter dopamine and projected into exactly one matrix neuron of the striatum, whereby the long path caused a time delay.
- On the one hand, this matrix neuron was tonically excited, since it tapped the axons of the cortical neurons passing through the striatum.
- At the same time it was inhibited by the incoming dopaminergic signal.
- This corresponded to a signal inversion. The output signal of the matrix neuron was therefore the inverted and to GABA switched cortex signal of the corresponding class 3 cortex neuron.
- The matrix axon reached an assigned neuron in the globus pallidus. This neuron was permanently excited by the influx of the mean value signals from the nucleus subthalamicus. This permanent excitation was inhibited by the matrix axon. This corresponded to a second signal inversion.
- So the output of the globus pallidus was the double inverted signal from the corresponding cortexneuron.
- Since a double signal inversion returns the original signal, the output of the neuron in the globus pallidus corresponded to the original cortex signal, but was inhibitory due to the transmitter switching to GABA.
- The basal ganglion system consisting of striatum, globus pallidus, substantia nigra pars compacta and nucleus subthalamicus caused the switching of the signal to the transmitter GABA for each class 3 cortex neuron, the two signal inversions that were carried out in this process cancelled each other out. However, the original cortex signal was delayed more on the long signal path.
Theorem of the extended basal ganglia system
The output of the globus pallidus was the time-delayed and switched to the inhibitory transmitter cortex output of class 3 neurons.
Now nothing more stood in the way of the formation of a time-sensitive difference mapping. This was because the cortex projected descending into the motor thalamus, where all class 3 signals were present in excitatory form.
If the globus pallidus projected back into this thalamus in a point-to-point image, then there was an excitatory signal in each thalamic neuron and the time-delayed and inhibitory signal obtained from it came from the globus pallidus. Thus the thalamus - wherever it was supplied simultaneously by the cortex and the globus pallidus - became a time-sensitive differential circuit and was able to detect movements and changes in signal strength. Its output was a result of the movement and change analysis.
And since these thalamus regions also projected towards the cortex, association areas could be created in the cortex to analyze movements and signal strength changes. Thus, the original movement analysis regarding signal class 5 in the nucleus ruber was supplemented by an additional one in the thalamus, which evaluated the signals of the divergent signal class 3.
Not all thalamus areas are used for motion analysis. The analysis of axon connections in the brain of vertebrates shows that the nucleus centromedianus and the nucleus ventralis (since time immemorial) serve for motion analysis.
Theorem of thalamic time-sensitive differential mapping for motion detection
In the nucleus centromedianus and the ventral thalamus, the excitatory thalamic input, which travels to the cortex, overlaps with the cortex signals returned from the globus pallidus, which are time-delayed and switched to GABA, in a point-to-point image, and generates a time-sensitive differential image that can analyze movements and signal strength changes. This thalamic output feeds the cortical association areas, which are responsible for the movement analysis of various modalities.
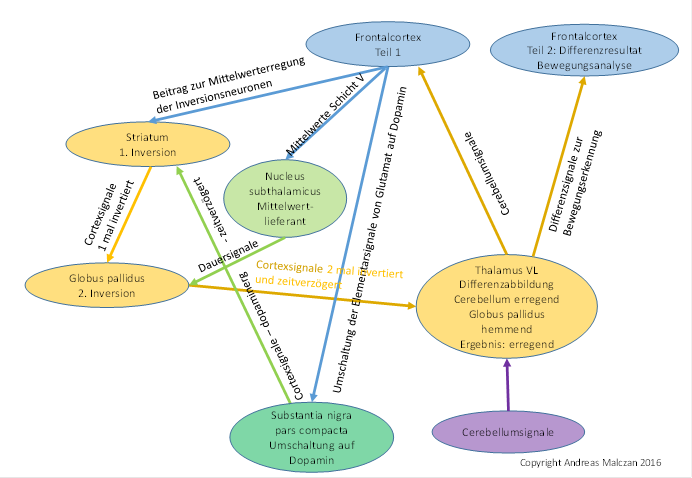
Figure 73- Signal inversion in the basal ganglia to generate a time-sensitive differential image in the thalamus VL
It should be noted that the Thalamus VL must be functionally divided into two parts. One half sends the output to the frontal cortex and the other half to the thalamus core. The second half receives the inhibitory signals of the globus pallidus and forms the difference image by point overlapping, which in turn reaches new association areas in the frontal cortex.
In higher vertebrates, the thalamic putamen is (presumably) responsible for optical motion analysis, while the ventral thalamus is more responsible for motor motion control.
As a precautionary measure, it should be noted that the globus pallidus should divide again in the context of a functional expansion of the cerebellum and from this would emerge two independent nuclei, which are called inner and outer pallidus. In this context also the 1:1 projection in the striatum was lost, so that (in the higher mammals) a neuron of the substantia nigra pars compacta projected into a whole chain of striate neurons. This development was still a long way off here. It began, however, with the output of the cerebellum finding its way into the cerebellum. This led to the development of the pontocerebellum, which subjected the cortical signals to a further evaluation algorithm and gave them the ability to develop above-average intelligence.
The compartmentalization of the striatum into the matrix and the striosomes had an unexpected side effect, which was however predictable from a signal theoretical point of view. The striosomes largely represented signal class 5, the matrix represented signal class 3, the latter resulting from strong signal divergence from class 5. Here, the signal divergence could be reversed if the matrix would again project onto the striosomes. Indirectly such an effect can be observed. In the matrix, the dopaminergic axons not only reach the matrix neurons, but also excite local interneurons that use the transmitter acetylcholine and project excitatory signals into the striosome neurons. This projection restores the signal convergence of signal class 3 to signal class 5.
The following figure illustrates the signal processing algorithms in the basal ganglia system and highlights in particular the divergence and convergence of the signals.
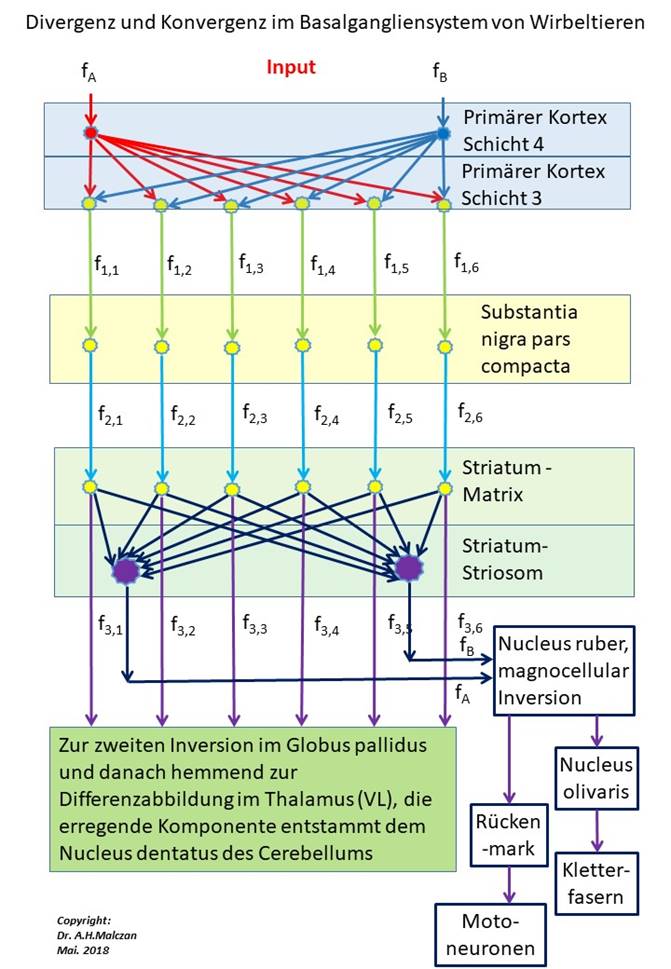
Figure 74- Divergence and convergence in the basal ganglia system
The class 3 dopaminergic axons that inhibit matrix neurons excite via collateral cholinergic interneurons that are distributed in the matrix space. These interneurons take over the excitation and in turn project excitation into the striosomal neurons. In this way, the original cortical signal divergence of class 5 is reversed to class 3 because the dopaminergic axons of class 3 of the matrix now project onto striosome class 5.
This projection is indirectly mediated by the cholinergic interneurons that possess the transmitter acetylcholine. Investigations of the structure of the striatum revealed compartmentalization at the unequal distribution of the transmitter ACH.
Theorem of signal convergence of the matrix signals to the striatum signals
The dopaminergic axons, which transport the signals of the class 3 cortex neurons to the matrix, excite cholinergic interneurons, which in turn excite striosomal neurons. Thus, the cortical signal divergence is reversed by striatal signal convergence and the class 5 signals are recovered from the class 3 signals in order to continue to supply the original time-sensitive differential mapping in the nucleus ruber with signals.
The original time-sensitive differential mapping in the nucleus ruber was thus preserved in the vertebrates, and they now also had a much more sensitive differential mapping in the ventral thalamus, which was based on the class 3 extreme value-coded signals obtained via divergence. This had consequences for the class 3, 5 and 6 signals, which were evaluated in the basal ganglion system. The signals derived from class 3 ended as inhibitory output of the globus pallidus in the ventral thalamus and overlapped there with the incoming excitation signals to form a time-sensitive differential image. The signals derived from class 5 also ended up in the nucleus ruber for the differential imaging there. The signals derived from the mean value signals of class 6 reached the nucleus ruber but did not find compatible partners there. They were transferred to the nucleus olivaris and arrived via the climbing fibers in the part of the cerebellum that developed into the pontocerebellum.
Since the output signals of this climbing fiber projection were derived from the cortical mean value neurons of class 6, which projected via the striatum into the nucleus ruber and to the contralateral cerebellum, we refer to this climbing fiber class as the mean value climbing fibers. It was discussed in more detail in the chapter on the Pontocerebellum.
Theorem of mean climbing fibres
We refer to the cortical projection of the mean value neurons of class 6 via the substantia nigra pars compacta to the striosomes of the striatum and from there via the nucleus ruber to the nucleus olivaris as the mean value projection into the climbing fiber system, the corresponding climbing fibers are called mean climbing fibers. They end in the Pontocerebellum at the Purkinje cells, but they also contact the cerebellar nuclei and the interneurons of the cerebellum.
This projection is responsible for the learning ability of the cerebellum, as shown in this monograph. In this context the synchronization of the different mean signals in the substantia nigra pars compacta is significant. The tetanic oscillations of the different mean values, which are interrupted by short pauses, are synchronized in such a way that the occurring pauses occur approximately simultaneously and have a frequency of 2 to 5 Hz. Per second there are therefore 2 to 5 pauses in the mean value oscillations, which are synchronised in time. This is significant for the functioning of the Pontocerebellum.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan