Vertebrate brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-064888-5
5.6 directional selectivity of the visuel divergence grating
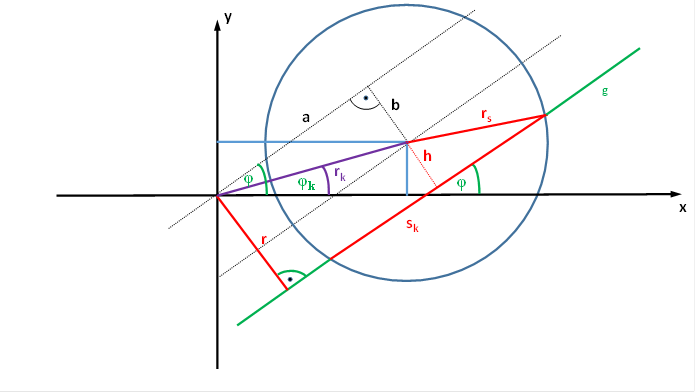 Figure61
- Chord length calculation for a shifted receptive field
Figure61
- Chord length calculation for a shifted receptive field
With reference to the above figure:
![]()
![]()
![]()
![]()
![]()
![]()
We assume that a dark on ganglion magnocellular cell, whose receptive field is intersected by a straight line with the angle of rise φ and the distance r from the coordinate origin, has a fire ratefk , which is a function of φ and r.
![]() (4.2.2)
(4.2.2)
We now investigate the firing rates f12, f2, f3 and f4 of four retinal ganglion cells of the described type under the assumption that they are arranged in the Cartesian coordinate system exactly along the coordinate axes and have a distance r = 1 from the coordinate origin. This is sketched in the following figure.
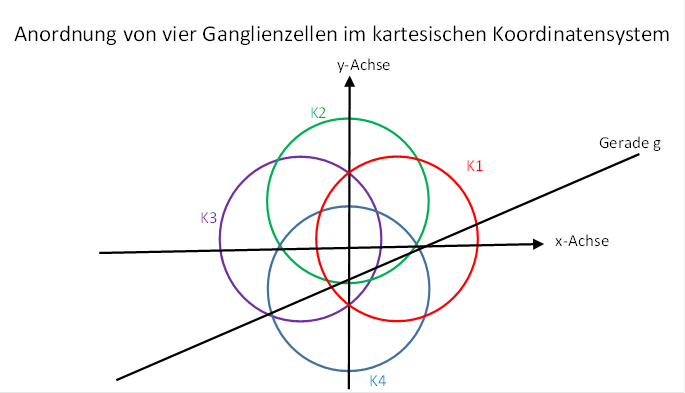
Figure62 - Arrangement of four visual ganglion cells
Each of the four ganglion cells has a circular receptive field with radiusrs.The partial or complete overlapping of the four receptive fields is important. The straight line has again the angle of ascent φ.
The following special features apply to this arrangement of ganglion cells:
![]()
![]()
![]()
![]()
![]()
This simplifies the formulas for the rate of fire
![]()
of the associated retinal ganglion cells:
![]() (4.2.3)
(4.2.3)
![]() (4.2.4)
(4.2.4)
![]() (4.2.5)
(4.2.5)
![]() (4.2.6)
(4.2.6)
These four fire rates travel from the retina via the Corpus geniculatum laterale to the visual cortex or visual cranial turning loop from which it emerged. All following considerations now refer to the signal propagation of these four input signals in the visual cortex. The topological arrangement of the four input signals should be preserved. Therefore there are four input neurons in the visual cortex, which in turn form the corners of a square and whose fire rates are the valuesf1, f2, f3 and f4according to our formulas.
We fill the square area with output neurons, which get their excitation via interneurons from the four input neurons. Simplified, we can divide the square by equally spaced horizontal and vertical grid lines, in whose intersections the output neurons may be located.We select any output neuron and calculate its excitation. This is made up of four parts, because it receives an excitation part from each of the four input neurons. However, a distance-dependent damping occurs, for which we assume a quadratic influence of the distance. The rate of fire of the considered output neuron therefore satisfies the equation
![]() (4.2.7)
(4.2.7)
The fire ratesfkoriginate from the described retinal ganglion cells.
We can also derive suitable formulas for the distancesr1, r2, r3 and r4of the output neuron to the four input neurons. For this we need the following figure.
We note that the input neurons lie on the coordinate axes according to our specifications and have the distance 1 from the coordinate origin.
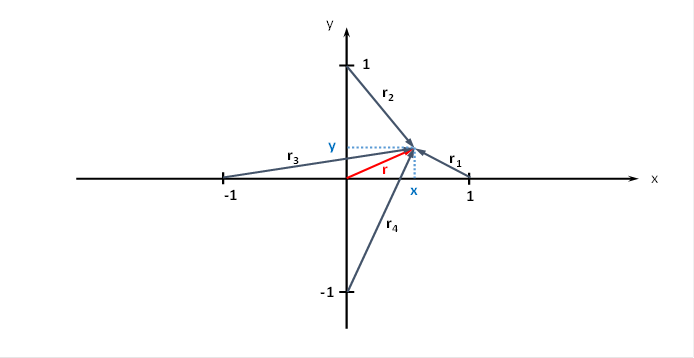
Figure63- Radius vectors to a neuron at point P(x,y)
At the latest here it becomes clear that in this case, too, we are dealing with a plane divergence grating, for whose excitation function we have already derived the formula (3.6)
![]() (4.2.8)
(4.2.8)
If there is an inclined straight line in the field of vision of the four retinal ganglion cells, we can insert the calculated fire rates into this equation and obtain for the total excitation f of the considered output neuron the equation
![]()
![]()
![]()
![]() (4.2.9)
(4.2.9)
We recapitulate
the most important requirements: The output neuron in the point![]() is
immovable. It cannot move actively.
Therefore the parameters
is
immovable. It cannot move actively.
Therefore the parameters![]() and
and ![]() constant
and unchangeable.
constant
and unchangeable.
Furthermore, we assume that the angle of rise φ of the straight line, which also overlaps the receptive fields of the four retinal ganglion cells, also remains constant.
If now the line is shifted parallel to itself, only its distance r from the coordinate origin changes. In this case it is observed in the experiment that the output neuron fires at maximum exactly when the angle of rise of the straight line takes on a very specific value. Neighboring output cells show a higher rate of fire, but at different angles.Mathematically,
this means that the fire rate f of the output cell under consideration in the
point![]() is
a function of the distance r and has a maximum. This means that the first
derivative from f to r must be zero, while the second derivative to r should be
negative and non-zero. Precisely then the excitation function f would have a
maximum. We therefore calculate the first and second derivatives to r. Here we
note that all quantities except r represent constants. We set the first
derivative to zero.
is
a function of the distance r and has a maximum. This means that the first
derivative from f to r must be zero, while the second derivative to r should be
negative and non-zero. Precisely then the excitation function f would have a
maximum. We therefore calculate the first and second derivatives to r. Here we
note that all quantities except r represent constants. We set the first
derivative to zero.
![]()
![]()
(4.2.10)
The second derivative after r gives the simple equation
![]() (4.2.11)
(4.2.11)
The equation resulting from zeroing the first derivative after r is first simplified by using the common factor
![]()
divide. This is allowed because this factor is not equal to zero. So we get the first simplification:
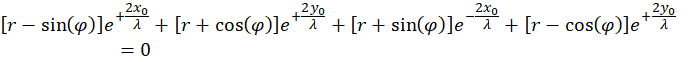
(4.2.12)
The conversion into the representation with hyperbolic functions is advantageous.
![]()
![]()
Using these relations we get the condition
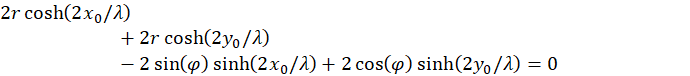
(4.2.13)
Thus we have a condition for
the existence of a maximum of the excitation function f of the output neuron in
the
point
![]() received:
received:
![]()
(4.2.14)
This equation of determination can be solved by the angle φ. For this we use the formula
![]()
![]()
![]()
we obtain the equation
![]()
![]()
(4.2.15)
This can be transformed into
![]()
(4.2.16)
Taking into account
![]() finally
results in the equation for the angle φ, where the output neuron is located at
finally
results in the equation for the angle φ, where the output neuron is located at
![]() isassuming
its maximum rate of fire f:
isassuming
its maximum rate of fire f:
![]()
(4.2.17)
![]() (4.2.18)
(4.2.18)
For this value
of φ the derivative of the rate of fire f to r is zero, the second derivative
being less than zero. Thus f assumes a local maximum. Thus, if the inclined line
separating the receptive fields of the four retinal ganglion cells involved has
this angle of rise φ and the distance r from the coordinate origin, the output
neuron fires in the visual cortex with the coordinat![]() and
has a local maximum. It is precisely this behavior that characterizes the
neurons of the orientation columns in the visual cortex. It is not based on
learning processes, but on the propagation of excitation of neighbouring
magnocellular ganglion cells in the visual cortex, whereby this propagation is
subject to a distance-dependent attenuation. In our model we postulated the
quadratic dependence of the rate of fire of the ganglion cells on the length of
the tendon, as well as the quadratic exponential damping. This model led
directly to an equation for determining the optimal angle at which the strongest
firing of the output neuron occurs. Other assumptions will lead to analogous
results.
and
has a local maximum. It is precisely this behavior that characterizes the
neurons of the orientation columns in the visual cortex. It is not based on
learning processes, but on the propagation of excitation of neighbouring
magnocellular ganglion cells in the visual cortex, whereby this propagation is
subject to a distance-dependent attenuation. In our model we postulated the
quadratic dependence of the rate of fire of the ganglion cells on the length of
the tendon, as well as the quadratic exponential damping. This model led
directly to an equation for determining the optimal angle at which the strongest
firing of the output neuron occurs. Other assumptions will lead to analogous
results.
We now use the possibility to represent the equation of determination as a function of x and y. Then, for example, we can display the relationship between the best angle and the location of an output neuron in Excel.
First of all,
the constant part in the diagram is
![]() of
the second summand is displayed graphically:
of
the second summand is displayed graphically:
Figure64 - The Angle Dependence of the Term T2
 The displayed angle was assigned a different color at intervals of 15
degrees (see legend in the diagram).
The displayed angle was assigned a different color at intervals of 15
degrees (see legend in the diagram).
This shows the contribution of the second summand in the equation of determination for φ. This contribution is independent of the distance r of the straight line to the coordinate origin.
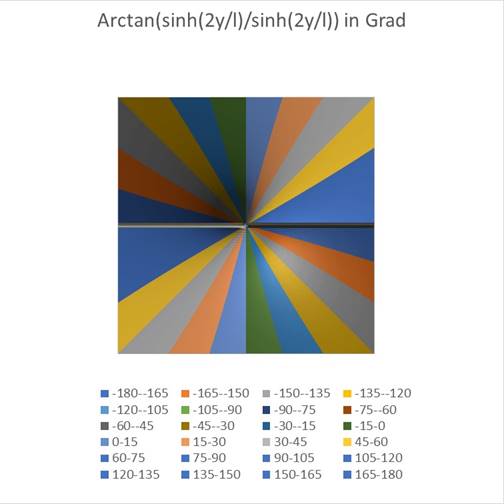
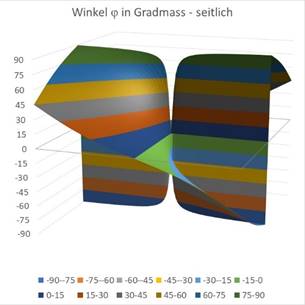
The following two diagrams show the angle φ of the total formula (18.5.3.21) as a function of the values x and y. In the left-hand diagram seen from the side and next to it from above. First we choose the value r = 0.
|
Figure65 - Display of the angle seen from the side |
igure66 - Viewing the angle from above |
The influence of the radius r can also be displayed graphically. In the above illustration r = 0 was given. The figures are Excel graphics, where the derived formulas for the angle φ were used to create the diagram. For a value of r = 0.2 the following graphs are obtained.
|
Figure67 - The influence of r on the directional selectivity |
Figure68 - The influence of r |
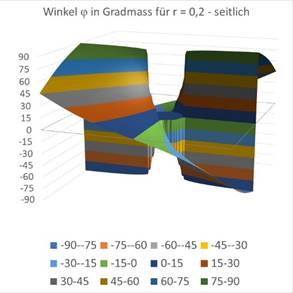
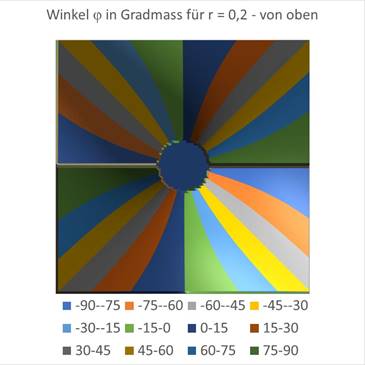
Here you can see that a circle-like small area of the coordinate origin responds to the angle φ = 0, while in the rest of the area the angles are arranged like a windmill, with each angle interval being assigned a different color in steps of 15 degrees. Exactly this was also done in the experiments on the angular selectivity of the orientation columns in the visual cortex V1. In this monograph, the mathematical context was presented for the first time.
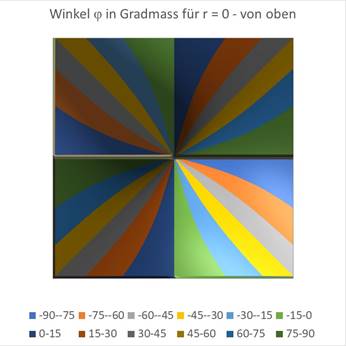
Finally the graphics for r = 0.5.
|
Figure69 - Orientation Columns for Large r |
Figure70 - Orientation columns with large r |
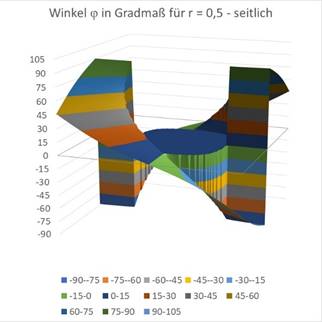
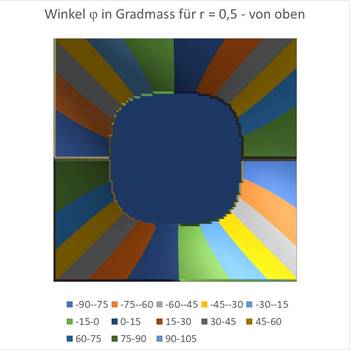
You can see that the circle around the coordinate origin, in which there is a maximum at an angle of incidence of 0 degrees, becomes larger as r grows. Approximately at the value of r = 0.7, the blue area is already so large that it fills almost the entire square. At about this value, the retinal ganglion cells no longer react with meaningful output because the inclined straight line no longer darkens the receptive fields sufficiently.
The blue circle at the origin of the coordinates, to which the tilt anglecalculable angle values - this is what the programmers from Excel decided. This solution is far better than issuing hundreds of error messages that would read: "Function value not defined!
herefore we have to prove at
this point why the function values are not defined in the "circle" shown in
blue. Equation (4.2.18) contains the term![]() .
.
![]()
An increase in the
value of the parameter r in the term ![]() &results
in the function's argument for certain parameter values being outside the
allowed interval. The greater r becomes, the greater the number of invalid
values.
&results
in the function's argument for certain parameter values being outside the
allowed interval. The greater r becomes, the greater the number of invalid
values.
The cause is simple: the length of the tendon must not be negative. If the inclined line no longer intersects the circle, there is no chord length and the formula leaves the permissible range.
To sum up, one could say
Theorem of the directional selectivity of orientation columns in the visual cortex
The directional selectivity of the orientation columns in the primary visual cortex and their mode of operation are based on signal propagation in plane divergence grids with distance-dependent, exponential signal attenuation. This signal divergence primarily increases the redundancy and thus the reliability of the system. Directional selectivity is the result of signal propagation on non-markless fibers where exponential attenuation occurs. The latter causes the directional selectivity.
By the way, this directional selectivity is independent of any movement - especially a parallel shift of the inclined line. Only the center distance r of the line to the coordinate origin enters the formulas, the angle of attack φ is constant. And it has been shown that for each angle of attack a different population of neurons fires at most, and these populations for different angles of attack are arranged around the center like a windmill, as can be seen in the diagrams.The special feature is that a flat divergence grating does not require any learning processes, its operating principles are present from the very beginning. Thus, the assumption that in the primary cortex fields - which do not receive their input from the spinocerebellum - learning neural networks are present proves to be unnecessary. The directional selectivity is not learned, it is present from the very beginning. It is based on elementary laws of nature, in particular the exponential damping of excitations when propagating in the surface.
This explains why nest prey are immediately able to detect objects. Objects are first recognized by the perception of their contours, which are detected as oriented line elements by the orientation columns in the cortex.
If the distance-dependent damping is not caused by an exponential function, but by another class of functions that is also convex, analogous results are obtained. In this respect, the function known as the cable equation can also be replaced by other functions.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan