Vertebrate brain theory
ISBN
978-3-00-064888-5
Monograph of Dr. rer. nat. Andreas Heinrich Malczan
5.5 The formation of divergence grids in the cortex
Motorized divergence grids for body monitoring and control
Plane convergence gratingsreverse the signal divergence caused by plane divergence gratings.
In the textbook "Neurosciences" [1] by Eric R. Kandel, James H. Schwartz and Thomas M. Jessell, Spektrum Akademischer Verlag 1995, chapter 29 presents the results of research on voluntary motor activity. It is shown that neurons in the primary motor cortex encode the force and direction of voluntary movements.
In an experimental arrangement, monkeys moved a rod with their arm to one of several target points, which were arranged in different directions around a central starting position.
During movement, the fire rates of a large number of cortical neurons were measured in the area where the signals of the receptors of the associated muscles and joints arrived.
It was shown that the direction of movement is not encoded by individual neurons, but by neuron populations. These neuron populations are arranged in a circle around a centre. Depending on the direction of movement, however, only one neuron population is active at any one time. Their position relative to the centre encodes the direction of movement in such a way that the population vector largely coincides with the direction of movement.
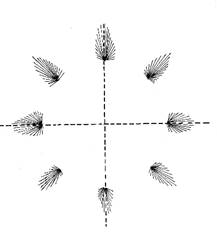
Figure 57 - Coding of the direction of motion neuron populations
Fig. 57 according to [1], page 545, Figure 29.3
And here we remember the excitation function in plane divergence grids. A joint like the shoulder joint allows the arm to rotate as desired. To simplify matters, one muscle group is responsible for lifting the arm forward. Another is responsible for lowering it. Both work against each other and provide the first pair of complementary signals. In these, the greatness u encodes the angle of the arm lifting forward.
Another muscle group is responsible for lifting the arm sideways, the corresponding antagonists lower the arm sideways again. Both work against each other again and provide the second pair of complementary signals. In these signals, the greatness v encodes the angle of the lateral arm lift. The interaction of the four muscle groups enables the arm to move in any (anatomically permissible) direction.
It is not difficult to assign a representation in polar coordinates to the prime quantities u and v. With
r² = u² + v²
and
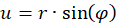
and
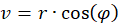
you get the radius vector r and the rotation angle φ. A circling arm movement would result in a circling radius vector r in polar coordinates. The larger the circle described by the circling arm movement, the larger the radius vector r in polar coordinates.
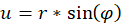 (4.1.1)
(4.1.1)
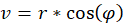 (4.1.2)
(4.1.2)
The corresponding muscle tension receptors of the four muscles (or muscle groups) involved transmit the excitations to the input neurons of the plane divergence grid in the motor cortex. The excitations overlap and have an excitation maximum, which rotates like the radius vector r when the arms are rotated. The length of the radius vector is determined by the size of the arm circles.
The excitation maximum at point P(xo, yo) is coupled to the magnitudes u and v by the following equations (3.14) and (3.15):
 (3.14)
(3.14)
 (3.15)
(3.15)
However, the neurons are not only excited at maximum excitation, but also in the immediate vicinity, even if the lateral inhibition ensures that more distant and therefore weakly excited neurons are inhibited more strongly and therefore hardly fire at all.
If the arm rotates, the excitation maximum and the surrounding active neuron population rotates around the coordinate origin. Whereby not the neurons rotate (they are stationary!), but the location with the strongest neuronal activity. And in this location there is not a single neuron, but a whole population of neurons. This real observed phenomenon of coding a direction of movement by a maximum coded neuron population proves that there are real existing divergence grids in the cortex in these areas, which behave as the derived theory predicts.
The special thing about this theory is that it does not require any learning processes. The explanation of the described phenomena using learning neural networks with plastic synapses can indeed provide similar results. But when comparing two different explanation theories, preference is often given to the theory that provides the simplest explanation. The action of ordinary laws of nature in the propagation of neuronal excitations on non-marrow fibres is quite sufficient to explain the phenomena in the motor cortex regions. Learning processes in neuronal networks are superfluous for this purpose.
And we should also bear in mind that such divergence grids are already present in the simplest creatures such as insects. They are the result of signal divergence - i.e. the transmission of neuronal signals on several axons. The output of such divergence grids allows for simple motor control - as observed in insect flight - without requiring higher brain power. Especially since some extremely short-lived insects would not have time to feed neural networks with huge amounts of data in order to learn movement patterns.
The reason for the extreme value coding is the non-linear signal attenuation on non-markless fibers. In this monograph we had made special assumptions about signal attenuation. However, it is quite possible to make other assumptions, to choose other functional equations. The results will be analogous. The location of extreme values will (almost) always be a continuous function of the function parameters, and the latter are the spatial coordinates of the neurons involved. One could therefore investigate which function classes lead to analogous results.
Theorem of maximum coding of directions of motion in plane divergence grids
In plane divergence grids in the motor cortex, the direction of movement is encoded by a maximum-coded population vector. This is caused by signal attenuation during the propagation of neuronal excitations.
It must be said that the activity of the neurons in the population vector is initially the result of the incoming receptor signals. Here, cause and effect must not be confused. The cause is the receptor signals and the associated muscle movements, the effect is the alignment of the population vector according to the direction of movement.
The situation would be different if other areas of the cerebral cortex were to send their signals into the flat divergence grid. This would also result in the formation of a population vector. In this case, however, the output of the divergence grid - after passing through a corresponding convergence grid - would be used for muscle control. Thus, thoughts could control the direction of movement.
The development of signal divergence in the primary cortex regions, which in the Bilateria's primordial brain were represented by the top level of the rope ladder system (which, as is well known, had split into different modalities), had far-reaching consequences.
The ascending axons from the neural tube or spinal cord reached the cortex via the thalamus of the associated modality (the second level of the original rope ladder system) and docked there to the class 4 neurons. These were now local interneurons. They transmitted their excitation to the class 3 neurons, which projected horizontally to the motorized ladder rail in the rope ladder system. As a result of the increasing signal divergence, the number of class 3 neurons increased enormously. The associated area expanded increasingly and formed a kind of spherical segment. When there was a lack of space, wrinkling occurred, which allowed the available area to be significantly increased. The associated axons became so numerous that they could no longer run horizontally to the motor ladder rail, especially since on the motor side there had to be an equally strong increase in class 5 receptor neurons. Thus, the outer surface of the cortex became spherical, and in its interior the axons of the class 3 neurons ran from the sensory to the motor part. There the necessary signal convergence occurred, special class 5 neurons (Betzsche pyramid cells) received the output of the corresponding divergence grating and cancelled the signal divergence. Thus, the neural tube or spinal cord received the number of motor output signals that had been significantly reduced by signal convergence.
The cortical divergence grids allowed a much better resolution of signals. Joint angles could be displayed and in the association areas could be combined much more finely with other, also more precisely resolved signals. The original yes-no principle, either fire or silence, was replaced by a much more precise evaluation of signals via maximum coded (or extreme value coded) signal populations. This allowed a much more finely controlled motor function, but was also a prerequisite for the development of higher intellectual performance. The extreme value coded signals resulting from signal divergence also reached the basal ganglia system and the pontocerebellum via class 5 neurons for further evaluation.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan