Vertebrate brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-064888-5
3.17 The topology of cortex and cerebellum clusters
For the analysis of the topology of the cortex and cerebellum clusters, we assume that the axons of the granule cells were already branching in a T-shape and aligned parallel to each other, so that they crossed the plane of the dendrite trees of the Purkinje cells at right angles. This assumes that the number of tactile and pain receptors was already significantly greater than the number of motoneurons, as already indicated at the end of chapter 3.15.
For the analysis of the signal projection from the cortex to the pontocerebellum we use the cluster model. A cluster (both in the cortex and in the cerebellum) is simplified as a square section of the cortex. Neighbouring clusters are about the same size. A cluster group consists of 9 clusters and is also square. The cluster with the number 5 is the inner cluster, the others are called outer clusters. The clusters 1, 3, 7 and 9 are the corner clusters. The direction in which clusters 4, 5 and 6 are arranged is called horizontal, they are arranged in a row.
We call the direction of clusters 2,5 and 8 vertical, these clusters form a column.
In the following figure a cluster group is shown, the 9 clusters are numbered as described.
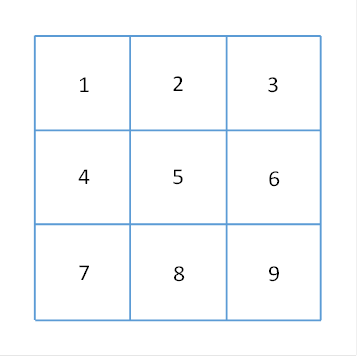
Figure 17 - Cluster group in the cortex
We refer to cluster 5 as the inner cluster and the others as the outer cluster or marginal cluster of the cluster group. We can assign a signal to each cluster, so the signal S5 belongs to cluster 5.
Within a cortex cluster, the signal neurons are distributed approximately equally. These are the cortical output neurons of class 5. The signal of such a neuron is called elementary signal. The following example shows a cluster with 25 output neurons, which are arranged in 5 rows of 5 neurons each. The real number of cluster neurons in the cortex is certainly many times higher in higher vertebrates. Here we are only concerned with the organizational principle of the cluster topology.

Figure 18 - Individual clusters in the cortex - schematic representation
It becomes clear that a cluster signal, such as S5, is composed of many elementary signals that can be arranged and indexed in columns and rows. If the output O5,2,4 belongs to cluster 5 in the second row and fourth column, and if the number 5 represents the number of rows and columns, then (very simplified) the signal S5 can be written as a matrix:
|
s5,1,1 |
s5,1,2 |
s5,1,3 |
s5,1,4 |
s5,1,5 |
|
s5,2,1 |
s5,2,2 |
s5,2,3 |
s5,2,4 |
s5,2,5 |
|
s5,3,1 |
s5,3,2 |
s5,3,3 |
s5,3,4 |
s5,3,5 |
|
s5,4,1 |
s5,4,2 |
s5,4,3 |
s5,4,4 |
s5,4,5 |
|
s5,5,1 |
s5,5,2 |
s5,5,3 |
s5,5,4 |
s5,5,5 |
=S5 (5.4.1)
Here, s5,k,l represents the elementary signal in the kth row and the lth column, which is supplied in cluster number 5 by the signal neuron in the kth row and the lth column.
We postulate a signal projection of the signal neurons of the cortex clusters via the moss fibers into the granule cells of the cerebellum clusters and assume that this signal mapping is topologically true in the sense that the cluster number is preserved.
Theorem of signal projection into the cerebellum clusters
The projection of the cortex signals via the moss fibre system to the granule cells is cluster-true.
But even within a cluster there is (normally) no mixing of the signals. The topology of the signal neurons described in the matrix (5.4.1) is transferred from the cortex cluster to the cerebellum cluster, so that the granule cells there are arranged in the same way. Thus the granule cells of the cerebellum form a body model of the cortical signal neurons of class 5.
The Purkinje cells receive the output of the granule cells and thus ultimately the cortex signals. Since this projection is cluster-true, a cerebellum cluster would theoretically only be able to receive the output of the corresponding cortex cluster if the axon branches of the granule cells were quite small and roundish. However, the special structure of the granule cell axons enables the Purkinje cell in the middle cluster to receive signals from cortex clusters 2, 5 and 8, because the axons of the granule cells are T-shaped and are aligned parallel to each other, so that in each cerebellum cluster, a Purkinje cell can also receive the signals of the cortex cluster above and below it. The special, extremely flat shape of the dendritic tree of Purkinje cells supports this possibility of also processing signals from above and below located cortex clusters. Without a benefit, this specification would certainly not have come about.
Theorem of vertical signal projection
As a result of the vertical signal projection via the longer, vertically aligned parallel fibres of the cerebellum, the Purkinje cell in a cluster of the cerebellum receives signals from the vertically adjacent clusters of the cortex. Here applies:
- Cerebellum cluster 4 receives signals from cortex clusters 1 and 7 via parallel fibres.
- Cerebellum cluster 5 receives signals from cortex clusters 2 and 8 via parallel fibres.
- Cerebellum cluster 6 receives signals from cortex clusters 3 and 9 via parallel fibers.
One could say that the middle cluster row of a cluster group also receives the cortex output from the cluster row above and below via the parallel fibers.
The parallel fibers ensure the vertical propagation of the cluster signals.
The transfer of excitation from the parallel fibres to the Purkinje cells is regulated by three principles.
Theorem of inner parallel fiber excitation of Purkinje cells
If the parallel fibre output originates from the middle cluster, excitatory synapses are formed between these parallel fibres and the Purkinje cells in the Purkinje cells of the middle cluster. Here, the number of parallel fibers to a cortical signal neuron is constant and has a constant synaptic strength, which we assign the value 1. We call this projection inner parallel fiber excitation.
The internal parallel fiber excitation ensures the excitation of the Purkinje cells of the central cluster by the cortex neurons of the associated central cluster.
Theorem of the input of the nucleus dentatus
Each Purkinje cell projects into an excitatory output neuron of the nucleus dentatus, this nuclear neuron additionally receives the excitatory output of the cortical cluster, which causes the inner parallel fiber excitation in this Purkinje cell, via collateral axons of the moss fibers. The Purkinje cell transmits the signal relationship with the signals of the middle cluster to the connected output neurons, and the strength of these signals in the Purkinje cell and the nuclear neuron of the nucleus dentatus is equal.
Note: This projection is not unique, several Purkinje cells can project into the same nuclear neurons. This will be explained later.
Thus - in relation to the signals of the middle cluster - the nuclear neurons of the nucleus dentatus do not provide an output signal, because the excitation of the Purkinje cell destroys the equally strong excitation of the connected nuclear neurons by inhibition.
Note: All nuclear neurons of a mid cluster receive the same signals via collaterals of the moss fibers.
Each excitatory nuclear neuron is also connected to an inhibitory projection neuron, which projects back to the nucleus olivaris. The function of this neuron will be explained later.
In relation to the output of the Pontocerebellum, a transformation took place at about this stage of development. In the early phase of the pontocerebellum, the supply of the opposite side with inverse mean value signals was in the foreground; later, temporal changes were taken into account by forming a difference map. As the number of signals increased, the mean value signal of a cortex cluster approached more and more to a tetanic signal, its rate of fire increased. The interruptions imposed by the striosome system led to sampled tetanic climbing fiber signals for each active cortex cluster.
The development of moss fibre projection led to the fact that an output neuron in the nucleus dentatus only responded with output if the assigned Purkinje cell received signals from both the inner and outer clusters. This presupposed that several clusters were active; only their joint activity led to output in the Pontocerebellum. This was the first step towards the recognition of complex signals.
Theorem of the output of the nucleus dentatus
Without the influence of signals from the outer clusters of a cluster group, the nuclear neurons of the nucleus dentatus connected to a Purkinje cell do not provide any output signal because the Purkinje cell and its nuclear neurons are equally excited, but the Purkinje cell inhibits the nuclear neuron and eliminates its excitation.
This zero signal output is particularly important in the initial phase of the pontocerebellum,as it is neutral in effect. Although it leads to resource consumption, which initially brings no benefit, it is neutral in its effects and above all not harmful. In the course of evolution this neutrality could even be transformed into a benefit. This is relatedto the lateral neighbourhood inhibition that is common in the nervous system of vertebrates.
It should be mentioned here that the Purkinje cells tended to form groups so that all group members projected into one and the same excitatory output neuron. Group formation is a kind of insurance against failure, it is caused by the golgi cells and will be explained later.
The cortical mean neurons of class 6 also possessed a lateral inhibition for contrast enhancement, which was caused by inhibiting interneurons. This lateral inhibition was transferred from the cortex cluster to the cerebellum cluster. In the cortex, each cluster inhibited all clusters in the surrounding area. Conversely, one could also say that each cluster received inhibitory excitation from neighbouring clusters.
Applied to the cerebellum, this means that each Purkinje cell in a cluster was inhibited by the signals from the neighbouring clusters. The inhibition neurons were GABAergic interneurons. When the dendrite trees grew larger and larger during evolution, but only grew in one direction, i.e. they were completely flat, and the axons of the granule cells also only grew in one - and in a right-angled - direction, the inhibitory interneurons inevitably split into two different classes, which are now known as star cells and basket cells.The direction of the parallel fiber axons is called vertical direction here, the growth direction of the dendrite trees of the Purkinje cells is called horizontal direction. For the vertical direction the star cells took over the neighboring inhibition, for the horizontal direction the basket cells. This is explained in more detail below.
In the following, we consider the excitation influx to a Purkinje cell located in the middle cluster and refer exclusively to signals from the outer clusters.
The excitation of a parallel fiber in the cerebellar middle cluster can also come from the cortex cluster located vertically above and below it. The cortex cluster 2 projects into the granule cells of cerebellum cluster 2, but its vertically oriented parallel fibers reach the middle cluster 5. Likewise, the excitation of the cortex cluster 8 reaches the cerebellar middle cluster 5. We call this projection a vertical projection.
Theorem of vertical star cell inhibition of Purkinje cells
The parallel fibers of the vertical projection transport the cortex output of the vertically adjacent cortex clusters into a cerebellum cluster. These signals are not cluster-specific. The Purkinje cells do not have direct synaptic contact with the parallel fibers of the cluster, but only via star cells. The signals of the vertical projection excite the star cells, which in turn are inhibited by the Purkinje cells.
The cortex clusters 2 and 8 project excitatory via the vertical projection into the star cells of the cerebellum cluster 5, these star cells inhibit the Purkinje cells there. The same applies to the projection of cortex clusters 1 and 7 into the cerebellum cluster 4 and to the projection of cortex clusters 3 and 9 into the cerebellum cluster 6.
The vertical star cell inhibition corresponds to the lateral inhibitionin the nervous system. Neighbouring clusters inhibit each other. Here, however, the star cells are only involved in vertical inhibition. The basket cells in the cerebellum have specialised in horizontal inhibition. In the cerebellum there is horizontal signal propagation via the basket cells.
Theorem of horizontal basket cell inhibition of Purkinje cells
Non-cluster signals from the left and right marginal clusters (1,4,7 and 3,6,9, respectively) reach the Purkinje cells indirectly via basket cells, which receive these signals from the parallel fibres of the marginal clusters, are excited by them and reach the Purkinje cells of the central cluster via long, horizontal axons in order to synaptically contact and inhibit Purkinje cells there.
It should be noted that edge clusters 4 and 6 are also reached by parallel fibers of clusters 1,7,3 and 9, because the parallel fibers are vertically oriented and have a considerable length. Therefore, the excitation of all edge clusters reaches the basket cells present in clusters 4 and 6, which in turn reach all Purkinje cells of the central cluster.
The horizontal basket cell inhibition corresponds to the lateral inhibitionin the nervous system. However, it only works in the horizontal direction in the basket cells.
A special feature of basket cell inhibition is that each basket cell forms such a long, horizontally running axon that all Purkinje cells of a horizontal cluster row are reached. In each of them, the axon forms collaterals that weave around the cell body of the Purkinje cell in a basket-like manner. In our early model of the cerebellum, a cerebellum cluster initially has only a single Purkinje cell. This should change later. In anticipation of this, we also assume that there can be several Purkinje cells in a cerebellum cluster.
All in all, it can be said that parallel fibres with signals from the same cortex cluster excite the Purkinje cells. Parallel fibres from foreign clusters excite the basket and star cells, whereby the basket cells are responsible for vertically adjacent clusters and the star cells for horizontally adjacent clusters, but both also receive signals from clusters above and below via parallel fibres. The star and basket cells in turn inhibit the Purkinje cells.
Thus, Purkinje cells are excited by signals from the same cortex cluster, but inhibited by signals from foreign clusters. This corresponds to the well-known lateral inhibition, foreign signals inhibit the output, here the outer clusters inhibitthe inner cluster.
Now we will examine the consequences of the lateral neighboring inhibition of the cerebellum clusters by the neighboring clusters. First we remember the theorem of the output of the nucleus dentatus. Without the influence of signals from the outer clusters, the output neurons of the nucleus dentatus are unexcited, i.e. neuronally completely inactive. Although the Purkinje cell is excited by the signals of the inner cluster, as well as the glutamatergic nuclear neuron in the dentate nucleus, the Purkinje cell inhibits this nuclear neuron. Since both are equally excited, excitation and inhibition cancel each other out. The output is the zero signal.
However, if signals from the outer clusters have an effect, the Purkinje cell is additionally inhibited by these signals with the interposition of the star and basket cells. This inhibition increases with the increasing signal strength of these signals. As a result, the remaining excitation of the Purkinje cell is no longer sufficient to completely inhibit the nuclear neuron. A residual excitation remains, which increases with the signal strength of the external signals. However, this is only true if the output neurons are excited by signals from the inner cluster, otherwise there is no output.
Theorem of the effect of the signals of the outer clusters on the output of the nucleus dentatus
With increasing signal strength of signals from the outer clusters, the neurons of the nucleus dentatus in the associated inner cluster will produce an output signal that increases with this signal strength, provided that the inner cluster also produces signals.
If the inner cluster does not generate any signals, the output of the nuclear neurons is the zero signal even if signals from the outer clusters are present.
This is mathematically a logical and - linking:
- Then, and only then, if active signals from the middle cluster 5 are present and additionally at least one outer cluster has active signals, the associated output neuron of the nucleus dentatus, which is synaptically linked to the signal-receiving Purkinje cell, delivers a (non-trivial) output signal.
This enables the circuit to react to combinations of signals. These signal combinations from different, neighbouring cortex clusters are usually referred to in neurology as complex signals.
First, a dentate neuron produces an output when any arbitrary, stronger signal from the inner cluster and simultaneously any arbitrary signal from at least one outer cluster arrives at the associated Purkinje cell. Through the effects of long-term depression (LTD) and long-term potentiation (LTP), a significant further development was to take place here, which gave vertebrates an ability to learn.
After this detailed derivation of the response reactions of Purkinje cells and cerebellar output neurons, we can offer a much shorter derivation of the same results, based on logical considerations and applying the ubiquitous principle of lateral inhibition.
To this end, we analyze the types of signal relationships between the different cerebellum neurons. They substantiate the previous findings.
The signal of the mean neuron of a cortex cluster is transmitted divergently on its way via the striosomes, the nucleus ruber and the nucleus olivaris from the climbing fiber axon to all Purkinje cells of the associated cerebellum cluster. The Purkinje cells therefore take over their signal relationship from the mean neuron of the cortex cluster and thus represent descendants of the mean neuron.
The signals of the signal neurons of a cortex cluster arrive as granule cell signals in the cerebellum cluster, therefore the granule cells of the cerebellum cluster are signal related to the signal neurons of the cortex cluster. In this respect they are descendants of the signal neurons of the cortex cluster.
Since the signal neurons of the cluster have a positive signal relationship with the mean neuron of the same cluster, they excite the mean neuron. This property is transferred to the offspring.
Therefore, the granule cells of a cerebellum cluster excite all Purkinje cells of the same cluster.
Thus, if the parallel fiber signal originates from the same cortex cluster as the climbing fiber signal derived from the mean neuron of the same cortex cluster, excitatory synapses are formed between these parallel fibers and the Purkinje cells.
Theorem of the origin of excitation of Purkinje cells
Signals from the same cortex cluster as the climbing fibre signal have an excitatory effect on Purkinje cells.
We can now make analogous considerations on the subject of competing signal relationships. They lead to the following result.
Theorem of the inhibition origin of Purkinje cells
Purkinje cells whose climbing fibre signal originates from the mean neuron of a cortex cluster are inhibited by signals from neighbouring clusters. The inhibition is caused by interneurons that are excited by these signals via parallel fibres. Neighboring clusters that lie in a vertical direction have an inhibitory effect on these Purkinje cells via star cells, the others via basket cells.
In this way, the basket and star cells take over the
lateral inhibition, which is passed on to them via the competing signals.
With the development of long-term depression and long-term potentiation, the response of the Pontocerebellum was specified. This is described in the following chapter.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan