Theorie der neuronalen Schaltung des Gehirns
und des analytischen Denkens
ISBN 978-3-00-037458-6 - ISBN 978-3-00-042153-2
Teil 2.9. Die Substitution des magnocellularen Kletterfasersignals durch das parvocellulare
Bisher haben wir viel Fleiß aufgewendet, um zu erklären, wie aus einem einzigen primären Kletterfasersignal, welches aus dem Aktivitätsneuron eines Cortexclusters durch Mittelwertbildung über den Signalneuronen des Cortexclusters hervorging, letztendlich viele hunderte oder tausende sekundäre Kletterfasersignale entstanden.
Hierbei wurde im einfachsten Falle das primäre Kletterfasersignal durch Verteilung auf die vielen sekundären Kletterfaseraxone aufgeteilt. Eine evolutionär erste Variante war die zentrale Verteilung über ein gemeinsames Verteilungsneuron im Nucleus olivaris.
Fortschrittlicher war die sequentielle Verteilung des primären Kletterfasersignals über eine sequentielle Verteilungskette. Man könnte meinen, nun hätte die Natur ihr Ziel erreicht. Wären da nicht die Unterschiede im Lernvermögen zwischen den verschiedenen Gattungen. Und gäbe es nicht das in dieser Monografie bisher ungelöste Problem der Intelligenzentstehung.
Wir haben erkannt, dass das Cerebellum zwangsgeprägt wird. Die Prägung erfolgt durch einmaliges Aktivwerden des Kletterfasersignals in einer freien Purkinjegruppe für die Dauer des Prägungszeitraumes. Damit ist aber der Lernprozess beendet. Die bisherige Theorie zeigt lediglich auf, wie im Cerebellum ein erstmals auftretendes Signal in einem sehr kurzem Prägungszeitraum von etwa einer Sekunde erlernt werden kann. Aber neben dem Neuerlernen gibt es auch die Modifizierung und Anpassung des bereits Erlernten an neue Gegebenheiten. Daher musste es eine neuronale Schaltung geben, die genau diese Fähigkeiten hervorbrachte. Diese zu erkennen war das Vorrecht der Mathematiker, denn die zugehörige Theorie ist eine mathematische. Die Theorie der neuronalen Netze lieferte den Algorithmus, bei dem Intelligenz entstehen kann. Grundlage ist ein länger andauernder Lernprozess. Hier haben wir einen zeitweiligen Widerspruch zur bisher vorgestellten Theorie.
Die Natur muss eine Möglichkeit gefunden haben, bereits erlerntes Wissen zu modifizieren. Und hier liegt der Schlüssel für den neuen Weg, den es offenbar geben musste.
Kein Lebewesen erlernt irgendetwas innerhalb von einer Sekunde und behält dieses Wissen auf immer und ewig. Genau das Gegenteil ist der Fall. Erworbenes Wissen wird ständig umgeschichtet, neu bewertet, aktualisiert. Wie also wird dieses Wissen, welches ja in den Purkinjezellen bereits fest eingebrannt ist, im Nachhinein modifiziert? Wie verläuft der ständige Lernprozess?
Wir versuchen, diese Frage in einen systemtheoretischen Kontext zu bringen und fragen anders:
Wie erfolgt die Nachprägung der Purkinjezellen?
Wenn ein starkes Kletterfasersignal die „Erstprägung“ bewirkte, wie könnte dann ein „neues“ Kletterfasersignal eine „Nachprägung“ bewirken, bei der das Wissen aktualisiert wird.
Beispiel:
- Erster Lernvorgang: Hunde fressen Knochen.
- Zweiter Lernvorgang: Wölfe fressen Knochen.
- Dritter Lernvorgang: Löwen fressen Knochen.
- usw.
- usw.
- Letzter Lernvorgang: Schakale fressen Knochen.
Wenn der erste Lernvorgang bei Fritzchen Müller am 01.März 2011 erfolgte, wie kann dann der zweite Lernvorgang am 30. Mai 2011 vor sich gehen. Und wie die anderen Lernvorgänge, wenn zwischen jedem einzelnen Tage, Wochen, Monate oder Jahre vergehen.
Die Purkinjezelle mit dem Inhalt „frisst Knochen“ muss auf den Input „Hunde“ reagieren, später zusätzlich auf den Input „Wölfe“, und letztlich auch auf den Input „Schakale“.
Mit jedem Lernvorgang kommen also neue Moosfasersignale als prägende Eigensignalanteile hinzu. Wie wird dies im Cerebellum gehandhabt.
Zunächst erscheint es aus Sicht des Autors dringend geboten, zur Verkürzung künftiger Texte und zur Schaffung einer gewissen systemtheoretischen Ordnung einige Begriffe zu definieren. Neurologen werden diese Termini bereits kennen. Hier jedoch mögen sie präzisiert werden.
Definition 2.7.: Primärer Cortex, primäre Cortexcluster
Die Cortexgebiete, die ihren Input von den Rezeptoren des Nervensystems erhalten, bezeichnen wir als primäre Cortexgebiete. Die zugehörigen Cortexcluster bezeichnen wir als primäre Cortexcluster.
Definition 2.8: Primärer (parvocellularer) Thalamus
Den Thalamus, der den primären Cortex mit den Rezeptorsignalen beliefert, nennen wir den primären (parvocellularen) Thalamus.
Da sich der primäre Cortex in Cluster einteilen lässt, können wir durch „Rückverfolgen“ des Inputs im primären Thalamus ebenfalls Thalamuscluster definieren.
Wir verwenden beim primären Thalamus die Präzisierung parvocellular, weil es noch einen primären Thalamus gibt, der die magnocellularen Signale zum Cortex sendet. Dies ist beispielsweise der Nucleus centromedianus thalami, evtl. trifft dies auch die intralaminaren Kerne zu. Wo also Verwechselungen möglich sind, sollte der Zusatz parvocellular verwendet werden.
Definition 2.9: Primäres Cerebellum, primäre Cerebellumcluster
Die Signalneuronen der primären Cortexcluster projizieren über die Brückenkerne in die Moosfasern und Kernneuronen eines Cerebellumclusters. Dieser Cerebellumcluster erhält zugleich den Kletterfaserinput, der vom Basalgangliensystem des gleichen Cortexclusters aus dem Aktivitätsneuron dieses Cortexclusters abgeleitet wurde. Hierbei ist es unwesentlich, ob die sekundären magnocellularen Kletterfasersignale durch zentrale oder sequentielle Verteilung entstanden. Diesen Cerebellumcluster bezeichnen wir als den primären Cerebellumcluster.
Im primären Cerebellumcluster entstehen also die Komplexsignale, die aus den Elementarsignalen des primären Cortexclusters zusammengesetzt sind.
Der Output des primären Cerebellumclusters erreicht über die positiven Kernneuronen den Thalamus. Auch dieses Thalamusgebiet erhält einen Namen.
Definition 2.10: Sekundärer (parvocellularer) Thalamus
Der Output der positiven Kernneuronen des primären Cerebellumclusters erreicht als erregendes Signal den sekundären (parvocellularen) Thalamus.
Der sekundäre (parvocellulare) Thalamus projiziert seinerseits in ein Cortexgebiet, welches wir als sekundären Cortex bezeichnen wollen.
Definition 2.11: Sekundärer Cortex, sekundäre Cortexcluster
Die positiven Kernneuronen jedes primären Cerebellumclusters projizieren über den zugeordneten sekundären (parvocellularen) Thalamuscluster in ein Cortexcluster des sekundären Cortex.
Die Signalneuronen der sekundären Cortexcluster wiederum projizieren über die ihnen zugeordneten Brückenkerne (Brückenkerncluster) in ein neues Cerebellumgebiet. Diesem geben wir ebenfalls einen Namen, um zukünftige Betrachtungen textlich kürzer fassen zu können.
Definition 2.12: Sekundäres Cerebellum, sekundäre Cerebellumcluster
Die Signalneuronen der sekundären Cortexcluster projizieren über die ihnen zugeordneten Brückenkernneuronen in ein Cerebellumgebiet, welches wir als den sekundären Cerebellumcluster bezeichnen.
Bekanntlich erzeugen (nach der Theorie des Autors) alle Cortexgebiete zweierlei Arten von Kletterfasern. Die magnocellularen Kletterfasern haben ihren Ursprung im Aktivitätsneuron des zugehörigen Cortexclusters. Das Striosomensystem der Basalganglien transformiert diesen Cluster-Erregungswert in ein magnocellulares, primäres Kletterfasersignal. Dieses wird über zentrale oder sequentielle Verteilung in die benötigten magnocellularen sekundären Kletterfasersignale umgewandelt. Daher hat jede Purkinjegruppe im primären Cerebellum-cluster ein eigenes sekundäres (aber noch magnocellulares) Kletterfasersignal.
Jedes im primären Cerebellumcluster erlernte (geprägte) Eigensignal besitzt eine unverwechselbare digitale Signatur. Die Dualziffer an einer k-ten Dualstelle hat den Wert 1, wenn die zugehörige Moosfaser des Cortexsignals zum Eigensignal der Purkinjegruppe gehört. Gehört das Signal der zugehörigen Moosfaser nicht zum Eigensignal, sondern zum Fremdsignal, so ist der Wert dieser Dualziffer gleich Null.
Wir können nun die miteinander verbundenen Neuronen des primären Cerebellumclusters, des sekundären (parvocellularen) Thalamusclusters und des sekundären Cortexclusters eindeutig beschreiben. Sie repräsentieren alle das gleiche Komplexsignal und besitzen die gleiche binäre Signatur.
Theorem 2.23: Signaturtheorem
Jedem gespeicherten Eigensignal des primären Cerebellumclusters entsprechen genau eine Purkinjegruppe, ein positives und ein negatives Kernneuron sowie genau ein Neuron im sekundären (parvocellularen) Thalamuscluster und im sekundären Cortexcluster mit der gleichen digitalen Signatur. Ist im zugehörigen primären Cortexcluster einen Auswahl von Signalneuronen hinreichend stark aktiv und entspricht dieser Neuronenaktivität ein Komplexsignal mit der gleichen digitalen Signatur, so erkennt (mit einer gewissen Zeitverzögerung) die Purkinjegruppe dieses Signal als ihr Eigensignal wieder, bewirkt die Erregung des zugehörigen Thalamusneurons und des zugehörigen Neurons im sekundären Cortex. Voraussetzung ist, dass das aktuelle Fremdsignal das Eigensignal nicht zu stark überdeckt.
Jedem Signalneuron im sekundären Cortex entspricht also ein Komplexsignal mit einer eigenen, digitalen Signatur.
Da aber jedes Signalneuron des sekundären Cortex ebenfalls in das Matrixsystem der Basalganglien projiziert, entsteht zu jedem dieser Signalneuronen genau ein parvocellulares Kletterfasersignal. Es ist seinerseits (mit einer gewissen Zeitverzögerung) immer genau dann aktiv, wenn das zugehörige Signalneuron im sekundären Cortex aktiv ist. Daher können wir diesem Kletterfasersignal genau dieses Komplexsignal zuordnen. Ebenso können wir diesem parvocellularen Kletterfasersignal genau die gleiche digitale Signatur zuordnen.
Definition 2.13: Digitale Signatur des parvocellularen Kletterfasersignals
Jedem parvocellularen Kletterfasersignal ordnen wir die gleiche digitale Signatur zu, die dasjenige Signalneuron im sekundären Cortex besitzt, von dem dieses Kletterfasersignal über das Matrixsystem abgeleitet wurde.
Nunmehr wird es etwas kompliziert, aber eine höchst wichtige Einsicht steht uns unmittelbar bevor.
Im primären Cerebellum gibt es zu jedem geprägten Komplexsignal eine Purkinjegruppe, deren digitale Signatur diesem Komplexsignal entspricht. Im Erkennungsfalle erregt das zugehörige positive Kernneuron genau ein Signalneuron im sekundären Cortex mit der gleichen Signatur. Dieses sendet über das Matrixsystem eine Kletterfaser los, die ebenfalls die gleiche Signatur besitzt. Alle diese Neuronen sind (mit geringer Zeitverschiebung) quasi synchron erregt.
Die Kletterfaser und ihr erzeugenden Neuron besitzen mit allen Neuronen der Signalkette eine ererbte Gemeinsamkeit. Sie erhielten von ihren Signalvorgängern, also ihren persönlichen Inputlieferanten, einen gemeinsamen Marker. Dieser ist eine Mischung verschiedenster Botenstoffe, die im System verteilt sind. Mindestens drei solcher Botenstoffe, die unabhängig sein sollten, ermöglichen eine Wegfindung im Raum. Dies liegt daran, dass jeder Botenstoff einen Ursprungsort besitzt, um den herum er sich z. B. durch Diffusion ausbreitet, wobei seine Konzentration abnimmt. Der Konzentrationsgradient ist eine Funktion der Entfernung vom Entstehungsort (oder der Entstehungsfläche).
Überlagern sich nun mehrere solcher Botenstoffe mit verschiedenen Entstehungsorten, so besitzt jeder Raumpunkt eine typische Botenstoffmischung. Diese markiert den betreffenden Raumpunkt und unterscheidet ihn dadurch von den anderen Raumpunkten (daher der Begriff „Marker“). Wird diese Botenstoffkombination vom Neuron an seine angeschlossenen Partner übergeben, so können diese am Ende der Kette den Weg zurück zum Ursprungsort zurückfinden. Dazu muss sich nur das zielsuchende Axon so im Raume bewegen, dass die ihm übergebene („vererbte“) Markerkonzentration als Suchgröße die Bewegung des Axons im Raume derart steuert, dass die extracellulare Markerkombination der internen möglichst entspricht.
Beispiele für solches Vorgehen werden in der Literatur beschrieben.
Das heißt, das Kletterfaseraxon kann seinen Ursprungspartner im primären Cerebellum finden. Der Autor dieser Monografie behauptet nun, dass jedes parvocellulare Kletterfasersignal genau dies tut.
Welche Gründe sollte es dafür geben?
Zunächst einmal muss festgestellt werden, dass die Purkinjegruppe, genauer ihr negatives Kernneuron, im Erkennungsfalle das zugehörige magnocellulare Kletterfasersignal hemmt.
Man vergegenwärtige sich die Hemmung an der Skizze 1.22 auf der Seiten 40. Und je mehr Purkinjegruppen eines Cerebellumclusters mit Eigensignalen geprägt worden sind, umso öfter wird auch eines dieser Signale nach seiner Prägung erkannt. Damit wird aber die Anzahl von Purkinjegruppen, die alle magnocellularen Kletterfasersignale hemmen, statistisch immer größer. Daher wird ein aktives Kletterfasersignal zum seltenen Ereignis, weil es immer seltener neue, bisher unbekannte Signale von hinreichender Intensität und Dauer gibt.
Dadurch wird der synaptische kontakt zwischen der Purkinjegruppe und dem zugehörigen magnocellularem Kletterfasersignal immer wirkungsloser.
Dies ist die große Stunde des parvocellularen Kletterfasersignals. Vom sekundären Cortex hervorgerufen, kommt es im Nucleus olivaris an und findet dasjenige Neuron, an dem das ursprüngliche magnocellulare Kletterfasersignal angedockt hatte. Diese kontaktstelle ist frei geworden, weil das magnocellulare Kletterfasersignal zu oft signallos und gehemmt war. Genau hier dockt das neue, das parvocellulare Kletterfasersignal an.
Dadurch wird die zugehörige Purkinjegruppe nunmehr (zusätzlich) vom eigenen, parvocellularen Kletterfasersignal versorgt. Beide Signale – das Outputsignal des Kernneurons und das parvocellulare Kletterfasersignal - haben die gleiche digitale Signatur. Immer, wenn der primäre Cortex über die Brückenkerne ein Komplexsignal ins primäre Cerebellum sendet, welches das Eigensignal dieser Purkinjegruppe ist, wird diese Purkinjegruppe (mit geringer Zeitverzögerung) vom parvocellularen Kletterfasersignal des zugehörigen sekundären Cortex erregt. Dieses ist das vom Autor lang gesuchte Phänomen der Nachprägung der Purkinjegruppen.
Jedes Mal, wenn dieses Komplexsignal im primären Cortex auftritt, wird die zugeordnete Purkinjegruppe im Cerebellum vermittels des parvocellularen Kletterfasersignals nachgeprägt. Sind während der Aktivität dieses Kletterfasersignals weitere Signal aktiv, die bisher nicht zum Eigensignal der Purkinjegruppe gehörten, so werden sie nun zusätzlich durch Langzeitpotenzierung und Langzeitdepression dem bisherigen Eigensignal hinzugefügt.
Dadurch wird das primäre Cerebellum lernfähiger. Nicht mehr die einmalige Prägung im Prägungszeitraum von etwa einer Sekunde ist die Ursache für das Gedächtnis, sondern die laufende Nachprägung mit wesentlichen Signalen.
Es muss jedoch darauf hingewiesen werden, dass der eben beschriebene Prozess der Nachprägung mathematisch höchst anspruchsvoll ist. Denn bei einer Kletterfaseraktivität werden ja per LTP neue Signale zum Eigensignal hinzugefügt, die im Prägungsmoment aktiv waren. Aber ebenso werden per LTD neue Signale zum Fremdsignal hinzugefügt, weil diese im Prägungsmoment gerade inaktiv waren. Das Resultat ist eine synaptische Kopplungsstärke, die nach dem Hebb‘schen Prinzip einer statistischen Häufigkeit dieses Signals entspricht. Die Eigensignalaktivität und die Fremdsignalaktivität eines jeden Bits der digitalen Signatur hängen vom statistischen Vorhandensein oder Nichtvorhandensein dieses Signals ab.
Außerdem hat das parvocellulare Kletterfasersignal nicht die gleiche Struktur wie das striosomale, magnocellulare Kletterfasersignal. Letzteres entspricht etwa dem neuronalen Systemtakt. Daher muss eine mathematisch und systemtheoretisch exakte Nachprägungstheorie erst noch vorgelegt werden. Hier wurde nur der systemtheoretische Algorithmus entwickelt. Mögen auch andere dieses fruchtbare Feld bestellen helfen.
Wir fassen unsere Erkenntnisse in ein neues Theorem. Wesentlichster Erkenntnispunkt ist die schrittweise Substitution der magnocellularen Kletterfasersignale durch die parvocellularen.
Theorem 2.24. Substitution des magnocellularen Kletterfasersignals durch das parvocellulare
Wenn eine Purkinjegruppe im primären Cerebellumcluster ihr Eigensignal erkennt, entfällt die starke Hemmung des zugeordneten dauererregten positiven und des negativen Kernneurons. Das negative Kernneuron hemmt im Erkennungsfalle das zugehörige Kletterfasersignal in der Olive, wodurch die synaptische Verbindung des zugehörigen Kletterfaserneurons zur Purkinjegruppe längerfristig wirkungslos wird, da die starke Kletterfasererregung immer genau dann gehemmt wird, wenn die Purkinjezellen wegen der Signalerkennung inaktiv sind. Der Output des positiven Kernneurons dieser Purkinjegruppe erreicht über den Thalamus ein Signalneuron in einem neuen sekundären Cortexgebiet. Dieses Signalneuron projiziert unter anderem in das Matrixsystem der Basalganglien und erzeugt ein neues parvocellulares Kletterfasersignal. Dieses parvocellulare Kletterfasersignal substituiert das inzwischen wirkungslose magnocellulare Kletterfasersignal und nimmt seinen bisherigen Platz ein. Da dieses parvocellulare Kletterfasersignal zwangsläufig immer genau dann aktiv ist, wenn die Purkinjegruppe ihr Eigensignal erkennt, wird genau in diesem Erkennungszeitraum eine Nachprägung der Purkinjegruppe mit dem parvocellularen Kletterfasersignal bewirkt. Die Substitution des magnocellularen Kletterfasersignals durch das parvocellulare erfolgt (wahrscheinlich) im Nucleus olivaris. Die Kernneuronen werden daher vom parvocellularen Kletterfasersignal miterregt, das positive Kernneuron meldet diese Erregung dem Thalamus.
Damit beschränkt sich der Lernvorgang im Pontocerebellum nicht mehr auf die Erstprägung der Purkinjegruppe. Bei jeder Signalerkennung erfolgt eine Nachprägung, so dass die Eigensignalanteile und die Fremdsignalanteile der Purkinjegruppe im Laufe der Zeit durch die Nachprägung aktualisiert werden.
In der nachfolgenden Skizze 2.7 ist das Prinzip der Kletterfasersubstitution dargestellt. Zu sehen ist der Zustand der neuronalen Schaltung nach der kompletten Substitution aller magnocellularen Kletterfasern des Striosomensystems durch die parvocellularen Kletterfasern, die von den Signalneuronen des sekundären Cortex über das Matrixsystem gebildet werden.
Skizze 2.7: Substitution der magnocellularen Kletterfasern des Striosomensystems durch die parvocellularen Kletterfasern des Matrixsystems
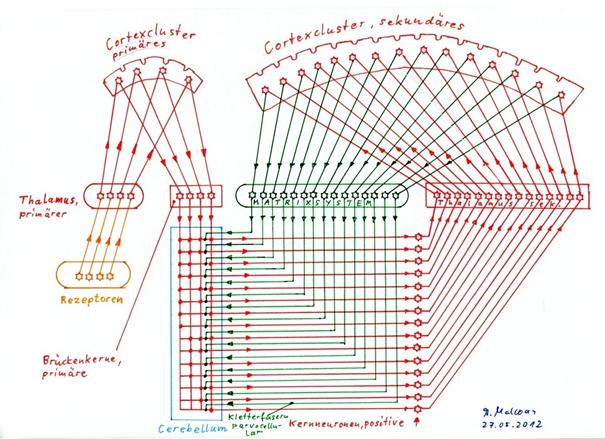
Das Matrixsystem, welches in der obigen Skizze als grüne Black-Box mit beispielhaft 15 Input- und Outputleitungen dargestellt ist, bildet nach Theorem 2.6 zu jedem Signalneuron eines Cortexclusters – hier des sekundären Cortexclusters – genau ein parvocellulares Kletterfasersignal durch vierfache Negation des Cortexsignals an einem Einssignal. Die in Skizze 2.2 dargestellte, relativ komplizierte Schaltung des Matrixsystems wird hier vereinfacht durch eine Box dargestellt, in der genau diese Signalumwandlung stattfindet. Erst dadurch kann die Kletterfasersubstitution, die den Übergang von Skizze 1.25 zur Skizze 2.7 beinhaltet, verständlich dargestellt werden.
Das Cerebellum ist in Skizze 2.7 vereinfacht dargestellt. Jedem Komplexsignal ist nur eine Purkinjezelle zugeordnet, Korb-, Stern- und Golgizellen sind weggelassen. Ebenso sind nur die positiven, erregenden Kernneuronen eingezeichnet, da sonst die Übersicht verloren gehen würde. Das Matrixsystem zum primären Cortex fehlt ebenfalls. Auch das komplette Striosomensystem wurde ebenfalls weggelassen.
An dieser Stelle erinnern wir uns an die Veränderung des Outputs der Kernneuronen während des Prägungsprozesses, die in Theorem 2.18 beschreiben wurde. Ist dieser Output bei Prägungsbeginn noch das Nullsignal, so stimmt er zum Prägungsende mit dem Output überein, den eine endgeprägte Purkinjegruppe erzeugt. Die der Nachprägung unterworfenen Purkinjegruppen sind bereits endgeprägt. Daher unterscheidet sich ihr Output während der Nachprägung nicht (wesentlich) vom Output in der Zeitspanne ohne Nachprägungssignal.
Dies ist eine überaus wichtige Erkenntnis. Da das durch die Kletterfasersubstitution an die (geprägte) Purkinjegruppe gelangende parvocellulare Kletterfasersignal (mit geringer Zeitverzögerung) immer dann aktiv ist, wenn die Purkinjegruppe ihr Eigensignal erkennt, darf dieses Kletterfasersignal die Rückmeldungen der Erkennung nicht beeinträchtigen.
Im Gegenteil: Es ist nunmehr völlig belanglos, ob die Purkinjegruppe ihr Eigensignal erkennt oder das Nachprägungs-Kletterfasersignal aktiv ist. Denn ein aktives Nachprägungs-Kletterfasersignal an der bereits geprägten Purkinjegruppe erhöht die Aktivität der als Eigensignaldetektoren wirkenden Sternzellen, Korbzellen und der Kernneuronen, aber auch die Aktivität der als Fremdsignaldetektoren wirkenden Purkinjezellen. Da aber die Korbzellen und die Sternzellen nun die stärker erregten Purkinjezellen stärker hemmen, ist die resultierende Erregung der Purkinjezellen nahezu unverändert.
Wir fassen unsere Erkenntnis in ein eigenes Theorem:
Theorem 2.25: Output der Kernneuronen geprägter Purkinjegruppen nach der Kletterfasersubstitution bei aktivem Kletterfasersignal
Nach der Kletterfasersubstitution bewirkt ein aktives parvocellulares Kletterfasersignal an einer geprägten Purkinjegruppe (in etwa) den gleichen Output wie die Erkennung des Eigensignals durch diese Purkinjegruppe. Das positive Kernneuron meldet mit seiner Erregung dem Thalamus die Signalerkennung, während das negative Kernneuron ebenfalls aktiviert ist und das magnocellulare Kletterfasersignal hemmt. Der Output während einer Nachprägung ist (fast) identisch mit dem Output während der Signalerkennung.
In etwa den gleichen Output möge interpretiert werden dahingehend, dass eine geringe Veränderung der Feuerrate möglich ist, aber die Erkennung nicht verändert.
Der Algorithmus der Nachprägung ist jedoch nicht nur mit einer realen Schaltungsänderung im Cerebellum verbunden, sondern auch mit einem Paradigmenwechsel.
Als Beispiel betrachten wir eine Purkinjegruppe vor der Substitution des magnocellularen Kletterfasersignals durch das parvocellulare. Wenn diese Purkinjegruppe ihr Eigensignal erkannt hatte, hemmte sie umgehend das magnocellulare Kletterfasersignal aus dem Striosomensystem in der Olive. Je nachdem, ob das magnocellulare Kletterfasersignal durch zentrale Verteilung oder durch sequentielle Verteilung gewonnen wurde, wurde das entsprechende Verteilungsneuron im Erkennungsfalle gehemmt. Diese Hemmung des magnocellularen Kletterfasersignals bleibt auch nach der Substitution erhalten. Durch die Beibehaltung der Hemmung des striosomalen Kletterfasersignals wird wie bisher verhindert, dass das gleiche Signal eine neue, freie Purkinjegruppe prägt. So werden Mehrfachprägungen auf die gleiche Weise wie bislang verhindert. Die Purkinjegruppe hemmt also weiterhin das striosomale Kletterfasersignal im Erkennungsfalle, auch wenn es nach der Kletterfasersubstitution gar nicht mehr von dieser striosomalen Kletterfaser erreicht wird.
Die Frage, ob das neue, parvocellulare Kletterfasersignal des Matrixsystems, welches nunmehr im Nucleus olivaris an das zuständige (bisherige) Kletterfaserneuron andockt, dort von der eigenen Purkinjegruppe (bzw. deren negativem Kernneuron) im Erkennungsfalle ebenso gehemmt wird wie bisher das magnocellulare, soll später beantwortet werden. Eine solche Hemmung erscheint widersinnig, denn beide sind (bis auf eine kleine Zeitverzögerung) synchron aktiv. Aber man darf die Phasenverschiebung der Signale auf Grund der längeren Ausbreitungswege nicht einfach vernachlässigen. Daher vertagen wir die Antwort auf später, wenn wir die Funktionsfähigkeit einer Schaltung diskutieren müssen, die eine Art inverser Videospeicher ist. Mit einer solchen Schaltung könnte das Kleinhirn vorher abgespeicherte Videosequenzen wieder so abspielen, z. B. während einer Traumphase.
Wir fassen die bisherigen Erkenntnisse in ein eigenes Theorem.
Theorem 2.26: Fortbestehen der Hemmung des striosomalen Kletterfasersignals nach der Kletterfasersubstitution
Nach der Substitution des magnocellularen, striosomalen Kletterfasersignals einer Purkinjegruppe im Nucleus olivaris durch das parvocellulare Kletterfasersignal des Matrixsystems bleibt die bestehende Hemmung des striosomalen Kletterfasersignals durch die Purkinjegruppe im Erkennungsfalle bestehen. So wird weiterhin eine Mehrfachprägung des Eigensignals dieser Purkinjegruppe im Cerebellumcluster durch das magnocellulare striosomale Kletterfasersignal verhindert.
Nach der Kletterfasersubstitution ändert sich prinzipiell nur wenig für das übrige Cerebellum. Das striosomale, magnocellulare Kletterfasersignal bleibt wie gehabt weiterhin zuständig für die Prägung neuer, bisher unbekannter Signale, wenn diese von ihrer Intensität und Dauer prägungsfähig sind.
Lediglich diejenigen Purkinjegruppen, bei denen die Kletterfasersubstitution von magnocellular auf parvocellular bereits erfolgte, sind zu einer Nachprägung fähig. Daher ist das Striomomensystem auch weiterhin für das Neuerlernen von Signalen im Cerebellum zuständig, das Matrixsystem jedoch für das Präzisieren und Aktualisieren des Gelernten.
Der Prozess der Nachprägung muss jedoch aus systemtheoretischer Sicht genauer analysiert werden.
Bei der Prägung veränderte sich die synaptische Kopplung der Eigensignaldetektoren durch Langzeitpotenzierung innerhalb von einer Sekunde vom Startwert ½ auf 1. Die synaptische Kopplung der Fremdsignaldetektoren veränderte sich durch Langzeitdepression in der Prägungssekunde von 1 auf ½.
Bei der Nachprägung geht alles deutlich langsamer und in kleineren Änderungsschritten. Der Autor vermutet, dass sich bei der Nachprägung der Kopplungswert nur im Prozentbereich ändern kann. Ist eine Parallelfaser im Cerebellum aktiv, die nicht zum Prägungssignal gehört, so wird durch das aktive parvocellulare Kletterfasersignal die Kopplung beim Eigensignaldetektor (z. B. Korbzelle) wie bisher vom Wert ½ auf den Wert 1 erhöht. Die bisherige Langzeitpotenzierung bleibt also unvermindert erhalten.
Wenn jedoch diese Parallelfaser zum Eigensignal gehört und während der Nachprägung inaktiv, also signallos war, muss sich der Kopplungswert wieder verringern. Diese Verringerung erfolgt jedoch nicht schlagartig von 1 auf ½, sondern wesentlich langsamer. Das Ersterlernen erfolgt also schnell, das Umlernen langsamer. Ein Beispiel möge dies veranschaulichen. Bettelt eine Katze oder ein Hund, während die Besitzer zu Tisch speisen, und das Tier erbettelt sich erstmalig einen Happen Futter, so wird es immer wieder um Futter betteln, während zu Tisch gegessen wird. Diese Bettelei abzugewöhnen, dauert viel länger. Der Erstlernvorgang „Betteln wird mit Futter belohnt“ ist sehr schwer umzuprägen zum Lernvorgang „Betteln wird nie mit Futter belohnt“.
Wir fassen diese Erkenntnis in ein eigenes Theorem.
Theorem 2.27: Schrittweise Änderung der synaptischen Kopplung bei der Nachprägung
Die durch Langzeitpotenzierung hervorgerufene starke Erhöhung der synaptischen Kopplung bei den Eigensignaldetektoren des Cerebellums kann durch eine Nachprägung nur schrittweise erniedrigt werden. Ebenso kann die durch Langzeitdepression bewirkte starke Erniedrigung der synaptischen Kopplung bei den Fremdsignaldetektoren des Cerebellums durch eine Nachprägung nur schrittweise wieder erhöht werden.
Die Umkehrung von LTP oder LTD wollen wir als inverse LTP oder inverse LTD auffassen. Hier kommen jedoch andere makromolekulare Vorgänge zur Anwendung, die von den Neurologen und anderen Forschern zunächst zu ermitteln sind. Der Autor vermutet lediglich, dass die inversen Prozesse deutlich langsamer ablaufen. Dies ist jedoch eine unbewiesene Hypothese. Es sollte bedacht werden, dass der Autor Mathematiker ist, also einen solchen Nachweis selbst nicht führen kann.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan