Theory of the brain’s neural circuitry
and analytical thinking
ISBN 978-3-00-037458-6 - ISBN 978-3-00-042153-2
Part 2.11. The basic circuit of the inner screen as inverse detection grid
An advantage of our thinking apparatus is the possibility to imagine something real or imagined. Dream experiences are particularly close to reality. Here we have the illusion that the dreamed is tangible reality. Only rarely do we recognize in a dream that we ourselves are dreaming. It would be the crowning glory of this monograph if the author succeeded in showing the circuitry of the "inner screen". Even if it were only a preliminary model.
Patience is recommended to the readers of this section. And all mathematicians and physicists among them are called upon to analyze the presented circuit from a systems theoretical point of view. The functionality is in the foreground.
Neurologists should critically but benevolently examine and question the circuit paths (projection paths) shown.
Although the associated scientific application is only envisaged in the third and fourth parts of this monograph, a brief insight into the author's theoretical concept will be given. The author of this monograph received his first real idea of how intelligent systems work many years ago when he first recognized how two cybernetic systems could communicate with each other. While one system sends out signals, the other system receives and interprets them. This system then responds while the other receives. The author preferred real language as a means of communication.
How does such a communication process work (in theory)?
First system A transmits and system B receives. System B converts the received sound into binary data. System B then speaks to system A. To do this, it first retrieves the buffered binary data, which is then transmitted as sound.
So there is a receiver circuit and a transmitter circuit. The author of this monograph realized at some point that these two circuits were inverse to each other. One of them converts sound into binary data, the other one transforms binary data into sound.
A mechanical analog without binary arithmetic was the invention of the record.
First, the sound moves a needle via a suitable membrane, which engraves the sound waves into a rotating wax disc, i.e. stores them. In the inverse process, the needle moves in the already engraved grooves of the rotating wax disc and moves a membrane which emits these movements as sound waves into the air. And to prevent the wax plate from wearing out so quickly, technicians thought of the possibility of transferring these grooves into more resistant plastic. The record was born. It is clear that the recording of sound and the playing of the record are inverse processes.
There are many applications of this kind in electronics. Radio transmitters convert sound into electromagnetic waves, which are converted back into sound waves by the radio receiver. In this way, an orchestra can simultaneously delight hundreds of thousands of listeners. We want to call such circuits inverse circuits.
According to the author, there is an inverse circuit in the brain to almost every neuronal circuit, in which the roles of input and output are reversed.
Some examples are given simply, although their explanation will be given in parts 3 and 4 of this monograph.
|
convergent circuit |
divergent circuit |
|
direct neural circuit |
inverse neural circuit |
|
Focus grid |
inverse center of gravity grid |
|
Interpolation grid |
inverse interpolation mesh |
|
Associative matrix |
inverse associative matrix |
|
Cortex |
inverse cortex |
|
Cerebellum |
inverse cerebellum |
The author is firmly convinced that there is an inverse circuit to almost every neuronal circuit, which also exists in the brain.
For example, there are cortex areas that receive their input from the thalamus and have a large number of output neurons as a divergent circuit to each input neuron. On the other hand, there are Cortexareals that have a relatively large number of input neurons but contain significantly fewer output neurons. The motor areas with Beetz giant cells belong to this category. He interprets these areas as inverse cortex. But more about this in parts 3 and 4 of this monograph, which have yet to be written. Here we concentrate on the theory of an inverse cerebellum.
If the video memory in the Cerebellum is a type of circuit, the author believes that there must be an inverse circuit. For example, this would be to play a film stored in the Cerebellum so that we see it in front of our inner screen. For example, during sleep, quasi as a dream experience.
That this inverse cerebellum has quite different real tasks to accomplish will probably become apparent in the third and fourth parts of the monograph. In particular, the entire motor function is based on the interaction of direct and inverse circuits, in which additional intelligence-giving neuronal networks and associative matrices are integrated. An inverse associative matrix should be of particular interest to mathematicians. The series connection of an associative matrix and an inverse associative matrix produces the original input as output. The series connection thus works as a single matrix, as a neutral element. Here, perhaps a new branch of algebra - signal algebra - is born.
So let us go to the explanation of the inverse cerebellum and the inner canvas.
We have recognized that primary and secondary cortex clusters can be distinguished. Each type of cluster produces parvocellular climbing fibers via the matrix system.
The parvocellular climbing fibers of the matrix system of the secondary cortex cluster couple to the corresponding Purkinj groups of the primary cerebellum and substitute there the magnocellular (striosomal) climbing fiber signal. This enables the primary cerebellum to be reimprinted. Learning processes become possible during the entire lifetime of the individual.
We now ask what is the function of the parvocellular climbing fibers, which are derived from the signal neurons of the primary cortex by the matrix system. To which Purkinj groups will these climbing fibres dock?
From a systems-theoretical point of view, the author of this monograph has only one possible variant: The parvocellular climbing fibers derived from the primary cortex cluster via the matrix system move to the secondary cerebellum. There they annex free Purkinj groups to imprint them with their climbing fibre activity.
These Purkinj groups are then marker-related to the signal neurons of the primary cortex. Each signal neuron transfers its personal, individual marker combination to the connected matrix neurons, which in turn create a parvocellular climbing fibre with this marker combination. The latter annexes a free Purkinje group in the secondary cerebellum and thereby also transfers the marker combination, i.e. the personal recognition code of the original signal neuron of the primary cortex, to the annexed Purkinje group. Therefore, the output of the Purkinje group can use the marker to search for the original signal neuron. The positive nuclear neuron projects into the thalamus and docks there exactly to the thalamus neuron whose signal leads to the original signal neuron. This completes the circle.
Thus there are two types of Purkinj groups in the secondary cerebellum. One species gets its parvocellular climbing fibers from the matrix system of the secondary cortex. According to the author, these Purkinje groups form the direct cerebellum. Their task is the reproduction of the Purkinje groups' own signals.
The other type of Purkinj groups get their parvocellular climbing fibers from the matrix system of the primary cortex. According to the author's theory they form the inverse cerebellum. We will first summarize this in a definition.
Definition 2.14: Direct and inverse cerebellum cluster
The direct cerebellum cluster is formed by those Purkinj groups that receive their moss fiber input from the primary cortex cluster with the involvement of the bridging nuclei, whose striosomal first imprint climbing fiber signal originates from the activity neuron of the primary cortex cluster and whose parvocellular post-imprint climbing fibers derive the matrix system from the signal neurons of the secondary cortex cluster. The associated cerebellum nucleus is called the direct cerebellum nucleus.
The inverse cerebellum cluster is formed by those Purkinj groups that receive their moss fibre input from the secondary cortex cluster with the involvement of the bridge nuclei and whose parvocellular climbing fibres are derived by the matrix system from the signal neurons of the primary cortex cluster. The associated cerebellum nucleus is called the inverse cerebellum nucleus.
Definition 2.15: Direct and inverse cerebellum
The entirety of the direct cerebellum clusters forms the direct cerebellum. The totality of the inverse cerebellum clusters forms the inverse cerebellum.
With this we have defined the term inverse cerebellum, but we have not yet proven that this is the neuronal circuitry of the inverse circuitry of the direct cerebellum. We will now provide this proof. Certainly, the proof could be simpler, more plausible or even aesthetically more beautiful. But we are not fighting here for a beauty prize, but for the accuracy of the proof.
The question is what claim we actually want to prove. Here the reader must rely on the author's intuition.
The author was inspired by the extremely exciting work "The Brain" by Richard F. Thompson from Spektrum Akademischer Verlag, which is also highly amusing to read, as shown below.
On page 266 we find the passage that first inspired the author to assume an inverse system in the brain whose task was to create illusions.
There we read the following anecdote about the visual-temporal field VTE:
(quotation begins:)
"The discovery that the VTE field receives a very specific type of visual information was a classic scientific stroke of luck. As the story is told, Charles Gross and his colleagues at Harvard University used monkeys to study the reactions of cells in the VTE field to visual stimuli. They used points of light, edges and bars - the usual standard stimuli. The nerve cells in the VTE region reacted a little bit to these simple stimuli, but not to touch or sound. Thus, the scientists thought that this region was a visual field, but not a very distinct one. After they had studied a certain cell for a long time with only minimal results - the cell hardly reacted to their stimuli - they decided to try another. As a gesture of farewell to this cell, one of the scientists raised his hand in front of the monkey's eye and waved. The cell suddenly fired like mad. Needless to say, the scientists stayed with this cell. They immediately cut out various hand outlines from paper and tried them out. The cell apparently liked a raised hand in the shape of a monkey's hand best. The cells in the VTE visual field seem to respond best to specific, complex outlines. Some neurones in this field react selectively to monkey faces, presumably in a similar way to the area that is responsible for the recognition of faces in humans.
(end of quote)
This example clearly shows how exciting and sometimes exhilarating neurological research can be.
But what does the shown selective ability of neurons in the VTE field - the visual-temporal field - have to do with the inverse cerebellum?
The author is firmly convinced that the mentioned "monkey hand neuron" from the above quote always fired strongly when the monkey saw a real monkey hand or at least a dummy of it in its visible field of vision. The scientists involved in the experiment could actually convince themselves of this.
But the author goes one step further. He assumes the claim that an artificial, external electronic stimulation of the "monkey hand neuron" by means of a small electrode would have given the monkey the illusion that a monkey hand had appeared in his field of vision.
The justification is simple, but must be proven.
It is already known that the electrical stimulation of neurons of the cortex or other regions is described by the test subjects involved in such a way that certain sensations, perceptions, even hallucinations occur. The author assumes that it is sufficient to electronically stimulate a neuron externally instead of confronting the nervous system with the real stimulus at which exactly this neuron fires. The neuron not only recognizes a complex signal assigned to it, but its external electronic stimulation creates the illusion in the nervous system that the complex signal is actually present.
It's similar in dreams. Here the illusion becomes perfect.
We want to present the neural circuitry for this and prove this thesis. This time we will proceed mathematically and put the assertion before the proof.
First, it seems reasonable to define the term complex neuron.
Definition 2.16: Complex neuron and its complex signal
We call every signal neuron of the secondary cortex a complex neuron. It is active exactly when the complex signal assigned to it acts on the primary cortex.
Let's recap: Each complex neuron is excited by exactly one neuron of the secondary thalamus. This thalamus neuron in turn receives its excitatory input signal from the positive nuclear neuron of the primary cerebellum exactly when the associated Purkinje group has recognized its own signal. This intrinsic signal is recognized when the associated complex signal acts on the primary cortex and its active signal neurons excite the moss fibers in the primary cerebellum that are part of the intrinsic signal of the Purkinje group. Thus, the own signal of the Purkinje group, the complex signal in the primary cortex and the complex signal assigned to the complex neuron are identical.
After these preliminary remarks, the proof must now be provided. The following assertion of the author must be proved:
(Beginning of claim:)
If a complex signal K which can be imprinted but has not been imprinted so far, consisting of k active elementary signals, comes from any receptors R1, R2, R3, ..., Rk of the system
a) in the primary thalamus the associated k neurons T1, T2, T3. ..., Tk excited,
b) these active thalamic neurons in turn excite the associated k neurons C1, C2, C3, ..., Ck in the primary cortex,
c) if exactly these cortexneurons k excite associated neurons B1, B2, B3, ..., Bk of the bridge nuclei,
d) if these bridging nucleus neurons in turn excite the moss fibres M1, M2, M3, ..., Mk of the associated cerebellum cluster
e) when these active moss fibres excite a free Purkinje group PGK in the cerebellum, while the climbing fibre signal derived from the activity neuron of the cortex cluster imprints this Purkinje group with exactly this complex signal K, so that its positive nuclear neuron in the secondary thalamus excites a complex neuron TS,K whenever the complex signal K is active
f) if this neuron of the secondary thalamus excites a neuron N of the secondary cortex when the complex signal K is active
g) when this neuron N of the secondary cortex activates a moss fibre MK of the secondary cerebellum in case of activity,
h) and if, in addition, each active signal neuron of the primary cortex C1, C2, C3, ..., Ck belonging to the complex signal generates a climbing fiber F1, F2, F3, ..., Fk via the matrix system of the basal ganglia, each of which has occupied and imprinted its own Purkinje cell P1, P2, P3, ..., Pk in the secondary cerebellum, which we call elementary signal Purkinje cells,
i) when each of these Purkinje cells P1, P2, P3, ..., Pk is activated by both the moss fibre MK and the associated climbing fibre F1, F2, F3, ..., Fk, so that imprinting occurs,
j) if the nuclear neurons K1, K2, K3, ..., Kk are assigned to these elementary signal Purkinje cells in the (inverse) secondary cerebellum nucleus,
k) when each of these elementary signaling Purkinje cells Ki in the primary thalamus contacts the neuron Ti and excites it in case of activity,
l) whenever the complex neuron N in the secondary cortex is externally stimulated via an electrode, but the complex signal K does not affect the receptors R1, R2, R3, ..., Rk of the nervous system
the primary thalamus is supplied, via the positive nuclear neurons K1, K2, K3, ..., Kk of the said elementary signal Purkinje cells, with the same complex signal K which it would have received if the real complex signal K had actually stimulated the receptors R1, R2, R3, ..., Rk of the nervous system.
(End of claim)
The external, electronic stimulation of the neuron N in the secondary cortex causes a moss fibre input to the elementary signal Purkinje cells. The output of their positive nuclear neurons provides the primary thalamus with the same input that an external complex signal would produce if it were present. Thus, the primary thalamus creates the illusion of a signal that does not exist in reality.
We now move on to the proof.
It may be that k active receptors send their signals as complex signals to the primary thalamus, where they excite k thalamic neurons. The receptor Rm might excite the thalamic neuron Tm.
Each thalamic neuron Tm projects into the primary cortex and excites a cortex neuron Cm.
Each of the k cortex neurons projects into a bridging nucleus neuron, whereby the cortex neuron Cm projects into the bridging nucleus neuron Bm and excites it.
Each bridging nucleus neuron projects into a moss fiber of the primary cerebellum, with the bridging nucleus neuron projecting Bm into the moss fiber Mm.
Since the complex signal K acting on the thalamus is imprintable and the imprinting has already been carried out, there is exactly one Purkinje group G in the primary cerebellum, whose own signal is the moss fibre signal arriving via the bridge nuclei. As long as K is active, the Purkinje group recognizes this signal.
The recognizing Purkinje group possesses a positive nuclear neuron, which is strongly excited in the case of recognition presented here and passes on its excitation via the secondary thalamus to a neuron N of the secondary cortex. Therefore, this cortex neuron N is also strongly excited as long as the complex signal K acts on the thalamus.
This cortexneuron N excites a moss fibre in the secondary cerebellum via the bridge nuclei, which in turn activates a parallel fibre pair (with a short time delay) whenever the complex signal K acts on the thalamus.
The k cortex neurons C1, C2, C3... , Ck of the primary cortex also send their input to the matrix system of the striatum. There, k parvocellular climbing fiber axons K1, K2, K3, ..., Kk are formed, which, according to the prerequisites, move to the secondary cerebellum and occupy k Purkinj groups there. Each of these climbing fibers is active (with a short time delay) exactly when the complex signal K of the primary cortex is active. However, this is exactly when the moss fibre mentioned above and the corresponding parallel fibre pair are also active.
Therefore, exactly k Purkinj groups are imprinted in the secondary cerebellum, if the complex signal K is active for a sufficient time for the first time. This is true, because this complex signal is imprintable according to the precondition.
Each of the k imprinted Purkinj groups receives a parallel fiber input from the parallel fiber population activated by the complex signal.
On the other hand, each of these Purkinje cells receives a climbing fiber signal, which is derived from the k signal neurons involved in the primary cortex. For this purpose, the output of these signal neurons, which are active according to the prerequisites, is converted into k active climbing fibre signals in the matrix system of the basal ganglia, which now characterise the Purkinje group mentioned above.
After this imprinting, each of these imprinted Purkinj groups will recognize its own signal exactly when the complex signal K is active in the thalamus and thus activates the neuron N in the secondary cortex so that its axon can excite the aforementioned parallel fiber population via the bridge nuclei.
However, the excitation of the parallel fibre flock is the reason for the Purkinj groups mentioned to recognise this signal and to excite the primary thalamus via their positive nuclear neurons. Here, the i-th nuclear neuron Ki excites the i-th thalamus neuron Ti.
If now the neuron N of the secondary cortex - as assumed - is not excited by the thalamus, but is stimulated by means of an external electrode, its excitation will also excite the same parallel fiber population via the bridge nuclei, which in the case of the Purkinj groups mentioned causes the signal recognition as an inherent signal. Therefore, the positive nuclear neurons of these Purkinj groups will excite the primary thalamus in the same way as the original real complex signal would have done. Except that in this case, one electrode will cause the stimulation of the neuron N.
The thalamus cannot distinguish who has excited it:
- Was it receptors R1, R2, R3, ..., Rk that excited him?
- Or were they the k output axons of the positive nuclear neurons of those elementary signal-Purkinje cells of the inverse cerebellum, whose climbing fibres came from the signal neurons T1, T2, T3, ..., Tk of the primary thalamus and whose moss fibre input came from the artificially irritated neuron N of the secondary cortex?
Of course, it does not necessarily have to be an external excitation using an electrode that applies a suitable excitation voltage to the neuron to be stimulated. Each neuron can also be stimulated by the higher-level systems. This is because the principle of internal mapping is recursive. From a systems theory perspective, it is based on the series connection of an associative matrix and an inverse associative matrix. Their output is the original signal, and according to the presented circuit it is fed to exactly those thalamus neurons which also receive the original signals. We summarize this knowledge in a separate theorem. It is a purely abstract information-theoretical theorem, which applies independently of the real neuronal circuit of the brain. In brackets are the real neurological references - rather for easier understanding. The author names this theorem after the city of Ulm, because the University of Ulm gave the first positive feedback on the first version of this monograph. However, the author is still waiting in vain for any reaction from Tübingen.
Theorem 2.28: Theorem of Ulm
If an input system (the thalamus) feeds its original signals to a circuit consisting of a series connection of an associative matrix (the direct cerebellum cluster) and an inverse associative matrix (the inverse cerebellum cluster), this circuit generates (for a certain input quantity) the original signals again as output. If the output of the series connection is fed back into the input system (the thalamus) as input, the system is not able to distinguish an external input into the inverse associative matrix from an original input into the input system. An external input to the inverse associative matrix makes the system believe that there is a real input to the input system (the thalamus) because the inverse system sends exactly one such input to the thalamus. The "inner canvas", the "inner eye of the observer" and the imagination and fantasy of humans and animals are based on this.
It must be clearly stated that Ulm's theorem needs to be worked on in greater depth. This is because associative matrices also react to input, which, in addition to the intrinsic signals to be recognised, may also contain external signal components. Therefore, the restrictive concept of "certain amount of input" needs to be specified more precisely through further research. For systems and information theorists, this is an uncharted field that needs to be worked on. Perhaps the University of Ulm has both the capacity and the initiative to take up and work on this topic.
Since the cerebellum represents an associative matrix, the author takes the liberty of calling the inverse cerebellum an inverse associative matrix, since it is also an associative matrix. However, this does not mean that the associative matrix and its inverse associative matrix are inverse to each other with respect to matrix addition or matrix multiplication. Inversity exists with respect to consecutive execution, which, however, is neither addition or subtraction nor multiplication or division. It will be shown later that the cerebellum represents a special case of an address decoder in terms of circuitry, while the inverse cerebellum represents a special address coder.
The author considers significant the possibility that the input into the inverse system does not necessarily have to be caused by external electronic stimulation of the neurons with electrodes. The higher systems, in turn, also have inverse components through which they can access the mentioned neurons.
Therefore, we must now ask ourselves why we do not constantly suffer from hallucinations, because some neurons of the secondary cortex are excited, for example, by superior subsystems. Here, the receptive neighbor inhibition of the thalamic neurons actively intervenes in the process. It is known that the thalamus contains a large number of inhibitory inter-neurons. Therefore, the stronger signals prevail. The output of the Purkinj groups of the inverse cerebellum is relatively weak, since only one parallel fibre population of many possible ones is excited. Therefore, the inverse signal generally falls through the excitation grid and is not particularly noticed.
Different in rest or low irritation. There suddenly "thoughts, images or even inner inspirations" appear out of nowhere. This is not a sign of illness, but the result of our inverse Cerebellum. It is able to transform signals from the higher spheres into our "inner view" or our "inner eye".
Therefore the following theorem applies.
Theorem 2.29: Receptive neighbor inhibition in the thalamus
The receptive neighbor inhibition in the thalamus is also necessary because the inverse system supplies the thalamus with input. It ensures that the strongest signals prevail over the weaker ones.
Chapter 3 will show that receptive neighbor inhibition - in which each output neuron inhibits each output neuron relative to its rate of fire - is a basic principle of all detection grids.
It should be mentioned that the receptive neighborhood inhibition does not only occur in the thalamus. There are inhibitory interneurons in all cerebellar nuclei, which apparently provide for receptive neighbourhood inhibition. Whether this receptive neighbourhood inhibition generally works between the output neurons, may be clarified by research.
Also in parts 3 and 4 of this monograph it is (probably) shown how the inverse cerebellum can be used to find answers to questions. Provided that someone is interested.
As usual, we summarize our findings on inverse cerebellum in a new theorem. As holder of the copyrights, the author decides to call exactly this theorem the theorem of Nowy Bytom in honour of his Polish birthplace.
Theorem 2.30: Theorem of Nowy Bytom
The inverse cerebellum as part of the secondary cerebellum is capable of transforming a natural or artificial excitation of a signal neuron in the secondary cortex, to which a complex signal K corresponds, back into the elementary signals of this signal in such a way that these excite exactly those signal neurons of the thalamus that would have caused a natural excitation of the neuronal system with this complex signal.
In contrast to the theorem of Ulm, which analyses the information-theoretical principle of the series connection of associative matrix and inverse associative matrix in a purely abstract way, the theorem of Nowy Bytom explains the concrete realisation in the (human) brain.
The sketch on the following page shows the basic circuit of the inner screen. On the left side the signal processing starts with the input that the receptors send to the primary thalamus. As an example, only 4 receptors are shown, although a cortex cluster is certainly supplied by some hundred input receptors.
The input rises to the primary cortex cluster and excites the four signal neurons that belong to it. The output of these neurons supplies two subsystems: the matrix system (shown on the left of the primary thalamus) and the bridge nuclei (shown on the right).
The bridge cores in turn supply the primary and direct cerebellum. The synapses of the 15 Purkinj groups are shown as dots, as well as the positive nuclear neurons. It would be too much to ask here to draw 15 complete Purkinj groups with, for example, three Purkinje cells each into this diagram, because then the overview would be completely lost.
With a little good will, mathematicians can already recognize the digital signature of the stored signals in the primary cerebellum. A bold cross point corresponds to a binary one, a missing one to the binary value zero. There are 15 non-trivial complex signals for four elementary signals of the primary thalamus.
The output of positive nuclear neurons moves to the secondary cortex. The output of the secondary cortex reaches the bridge nuclei and four corresponding Purkinj groups in the inverse cerebellum.
The climbing fibers for these four Purkinj groups come from the matrix system of the primary cortex (shown in blue). The inverse Cerebellum is inversely imprinted compared to the direct Cerebellum. Input are the 15 complex signals, output the four elementary signals.
If an external signal (as shown) stimulates a complex neuron of the secondary cortex, the secondary cerebellum transforms this input back into its original elementary signals and sends them to the primary thalamus. There the illusion is created that the complex signal is really present. In reality, however, only one neuron of the secondary cortex is stimulated.
While in sketch 2.8 on the following page the matrix system of the primary cortex cluster is drawn and sends its climbing fibres to the secondary, inverse cerebellum, the complete matrix system of the secondary cortex cluster with its 15 (example value) climbing fibres, which serve for the re-embossing in the primary cerebellum, is not drawn for reasons of clarity.
Sketch 2.8: The creation of illusions - The inverse cerebellum - The inner canvas
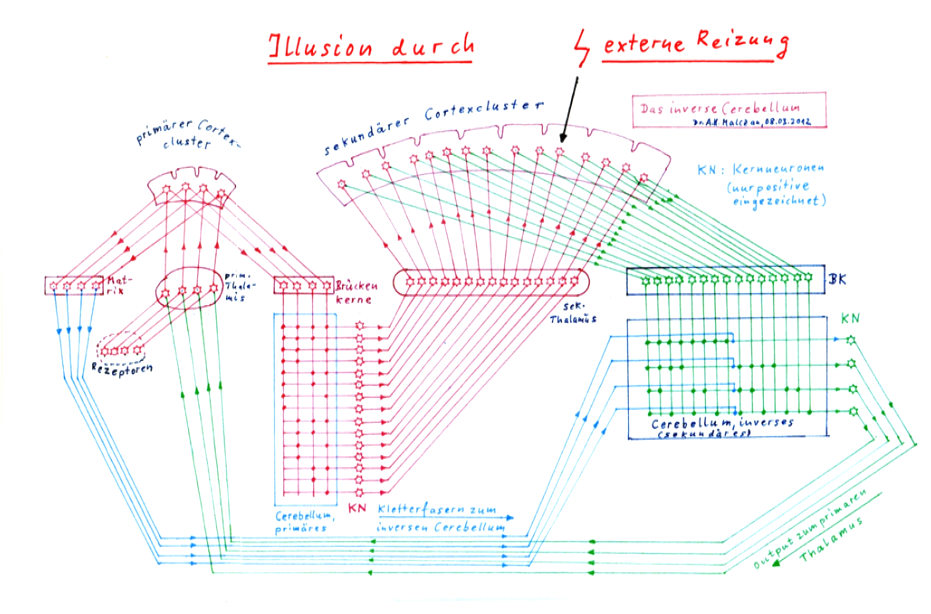
Thus, this theory divides the cerebellum into two partial cerebelli depending on the origin of the climbing fibres: the direct cerebellum and the inverse cerebellum. Likewise, two cerebellar nuclei are now distinguished.
While in the direct cerebellum the complex signals are stored, in the inverse cerebellum the elementary signals are stored. Since there are always many more complex signals to the elementary signals of the primary cortex, the direct cerebellum has considerably more Purkinj groups than the inverse cerebellum.
Each elementary signal can be active or passive, i.e. represented by one or zero. For two elementary signals there are already four combinations: 00, 01, 10 and 11. For 4 elementary signals there are theoretically 16 complex signals, for 5 elementary signals already 32. For n elementary signals there are 2n complex signals. At least in theory, because not every combination of elementary signals occurs in real life.
If all signal combinations were possible, an inverse cerebellum of n Purkinje groups would be assigned a direct cerebellum with 2n Purkinje groups.
Theorem 2.31: Potentiation theorem
For n elementary signals of the inverse cerebellum there are a maximum of 2n possible complex signals, if each elementary signal can only take the values active or inactive.
Here it is not yet considered that elementary signals could also be classified according to their activity, i.e. fire rate, e.g. by forming fire rate intervals.
The classification of analog fire rates into fire rate intervals is a major task of the brain's digitizing circuits, which (according to the author) are mainly realized by the cell columns of the cortex cortex. For more details, see the currently unpublished parts 3 and 4 of this monograph, which is planned to be published as an independent work.
Each elementary signal corresponds to a Purkinje group in the inverse Cerebellum. Likewise, each complex signal in the direct cerebellum also corresponds to a Purkinje group. While we could imagine the Purkinj groups of the inverse cerebellum lined up on a straight line, the Purkinj groups of the direct cerebellum would have to be lined up on an extremely long line due to their enormous number. Due to the potentiation theorem, this would be so enormously long that we would never be able to accommodate them as a straight line in our human body. However, nature has a good solution for lack of space. The straight line is bent and folded. This is how the large surfaces in the intestine or lungs are created. In the Cerebellum it is analogous. Due to the strong folding (wave crests and wave valleys) a line (or surface) with the Purkinj groups is compressed so far that it surrounds the inverse Cerebellum on the outside. On the inside - slightly elongated - lies the inverse cerebellum (worm). On the outside are the strongly folded surfaces of the direct cerebellum, almost infinitely finely folded and therefore unfolds a huge surface.
Theorem 2.32: Folding theorem of the direct cerebellum
The huge number of Purkinj groups of the direct cerebellum can only be accommodated in the cerebellar cortex because this area is accommodated by extreme and repeated folding in the relatively small space available. The inverse cerebellum, in contrast, forms a relatively straight body in the middle of this paired organ.
Likewise, the cerebellar nucleus of the inverse cerebellum consisting of n positive and n negative nuclear neurons would have its counterpart in a direct cerebellar nucleus consisting of 2n positive and negative nuclear neurons. The olive is also divided into a direct olive with a large number of neurons and an inverse olive with significantly fewer neurons.
If we consider that there are both ascending signals - the afferent signals - and descending signals - the efferent signals - in the brain, and that each of these signal types has its own direct and inverse cerebellum, we arrive at four cerebelli and four cerebellar nuclei.
This theory is not as far-fetched as it might seem. Neurologists actually distinguish four cerebellar nuclei in humans and higher mammals: the nucleus fastegii, the nucleus globosus, the nucleus emboliformis and the nucleus dentatus.
Theorem 2.33: Theorem of the four cerebellar nuclei
Since there is an afferent and an efferent system in the brain and each of them uses a direct and an inverse cerebellum, there are also four cerebellar nuclei: the nucleus fastegii, the nucleus globosus, the nucleus emboliformis and the nucleus dentatus.
Since the cerebellum is a paired organ, the number of its sub-elements doubles, with one sub-element being responsible for the left half of the body, the other for the right half, while in the worm both parts are represented.
It should be noted that cortex and cerebellum lumaxones cross each other at the transition from cortex to cerebellum and vice versa.
Since, in the author's view, the afferent system is recursively constructed because the output of stage 1 is used as input for another, afferent stage 2, whose output feeds the next higher stage and this recursivity is repeated, the nucleus dentatus, as the direct cerebellar nucleus of the afferent system, possesses an almost infinite number of nuclear neurons, the associated afferent direct cerebellum has an equally unbelievably large number of Purkinj groups and an almost perfect folding, in order to accommodate the growing number of complex signals with powers of two in a planar cerebellum bark.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan