Gehirntheorie der Wirbeltiere
für das Human Brain Projekt der Europäischen Union
ISBN 978-3-00-064888-5
3.22 Das analoge Urhirn der frühen Chordaten
Das bisher beschriebene Urhirnder frühen Chordaten realisierte seine gesamte Signalverarbeitung analog. Stets war die Feuerratedie maßgebliche analoge Größe. Dies traf einerseits zu bei der Transformation sensorischer Signale in motorische Steuersignale, wie es etwa im Torus semicircularisoder im Tectum opticumder Fall war.
Ebenso wurde die analoge Arbeitsweise bei der anfänglichen kontralateralen Hemmungund später bei der inversen Erregung der motorischen Gegenspielerrealisiert. Das Vestibulo- und das (frühe) Spinocerebellumarbeiteten analog über die Feuerrate.
Auch bei der Bewegungserkennung im Basalgangliensystemund in der Amygdalawar die analoge Feuerratedie maßgebliche Größe.
Selbst das urtümliche Pontocerebellum begründete seine Lernfähigkeit ausschließlich mit analogen Signalen.
Daher können wir zu Recht sagen, dass das frühe Urhirn der Chordaten analog arbeitete. Wir werden für das analoge Urhirnauch die Bezeichnung primäres Wirbeltiergehirnverwenden.
Theorem des analogen Urhirns
Das frühe Urhirn der Chordaten arbeitete analog.
Diese Aussage macht jedoch nur dann einen Sinn, wenn es weitere Arbeitsweisen des Chordatengehirnsgibt, die nicht analog erfolgen. Hier muss - auch gegen den erbitterten Widerstand etlicher Experten - die Arbeitshypothese vertreten werden, dass es im Verlauf der weiteren Evolution zur Herausbildung zweier weiterer Arbeitsweisen im Wirbeltiergehirnkam. Dadurch bildeten sich neue Substrukturen heraus, die man organisatorisch zusammenfassen kann. Letztlich ist das Wirbeltiergehirn dreiteilig gegliedertin ein primäres Subhirn, ein sekundäres Subhirnund ein tertiäres Subhirn. Das primäre Subhirn verwendet analoge Signale, das sekundäre bereits digitaleund das tertiäre Subhirn komplexe Signale. Und da der Autor diese Behauptung schrittweise beweist, müssen die Kritiker sich die Mühe machen und gravierende Fehler in dieser vorgelegten Beweisführung nachweisen. Es ist nicht hinnehmbar, dass man diese Theorie einfach nicht zur Kenntnis nimmt, um sie nicht widerlegen zu müssen! Insofern sind die Experten des Human Brain Projekts der Europäischen Union moralisch verpflichtet, diese Gehirntheorie der Wirbeltiere zur Kenntnis zu nehmen und zu prüfen.
Das Wirbeltiergehirn ist - funktionell, aber nicht räumlich - dreiteilig gegliedertin ein primäres Subhirn, ein sekundäres Subhirnund ein tertiäres Subhirn. Das primäre Subhirn verwendet analoge Signale, das sekundäre bereits digitaleund das tertiäre Subhirn komplexe Signale. Das primäre Subhirn ist räumlich diffus im sekundären Subhirn verteilt und bildet nur gelegentlich eigenständige räumliche Substrukturen wie etwa die Striosomen in der Matrix, den magnocellularen Anteil im Nucleus ruber oder die Betzschen Pyramidenzellen im motorischen Cortex. Es ist quasi versteckt und fällt nur bei sehr frühen Formen der Wirbeltiere auf. Das tertiäre Subhirn ist beim Homo sapiens voll ausgebildet, bei anderen Spezies weniger ausgeprägt. Seine Anfänge reichen dennoch in die frühe Urzeit zurück.
Jede dieser drei Strukturen verfügt über eine eigenständige Signalform. Jede der drei Strukturen tauscht ihre Signale mit den beiden anderen Subsystemen aus. Hierbei muss zuvor die bisherige Signalform (analog, digital, komplex) in die neue Signalform des Empfängersystems transformiert werden. Diese Signaltransformationen nehmen Transformationssystemevor. Zu ihnen gehören u. a. das Cerebellum, aber auch Teile des sensorischen Cortex, ebenso auch die Basalganglienund das limbische System.
Ebensolche Transformationsstrukturenmüssen durchlaufen werden, wenn die Signale eines Subsystems absteigend an die anderen Teilsysteme weitergegeben werden. Daher gibt es aufsteigend und absteigend projizierende Transformationssysteme. Zu den absteigend projizierenden gehört u. a. der Nucleus ruber, aber auch der motorische Cortexoder der dorsale ventrikuläre Kamm (DVR) bei Reptilien bzw. Vögeln.
Jede der drei Strukturen verfügt über ein eigenes Kurzzeitgedächtnisin Form eines Laufzeitsystems, bei welchem den Signalen eine kleine Zeitverzögerungaufgezwungen wird, während sie auf marklosen Axonentransportiert werden. Dieses Laufzeitsystem ermöglicht den Signalvergleich zwischen unverzögerten und verzögerten Signalen in Form einer Differenzabbildung, die vor allem der Bewegungserkennungdient.
Weiterhin verfügt jede der drei Strukturen über einen separaten Rotationsspeicher, in dem die Signale dauerhaft rotieren und so die Erinnerung an die Vergangenheit wachhalten, die bereits länger zurückliegt. Periodisch alle 24 Stunden - bei Bedarf auch früher - können die rotierenden Signale gelöscht werden. Hier ist der Nucleus suprachiasmaticuseingebunden sowie andere zeitgesteuerte Taktgeber.
Das tertiäre Systemverfügt über einen Komplexsignalspeicher, der wichtige Signale speichern und später wiedererkennenkann. Hier gibt es einen neuronalen Speicherbefehlsowie als Output ein Erkennungssignal, falls ein Signal erkannt wird.
Der Output des tertiären Systems hat wieder das analoge Datenformat und wird rekursiv wieder ins primäre System eingespeist. Dies wird in dieser Monografie nicht mehr behandelt, der interessierte Leser sei auf frühere Monografien des Autors hingewiesen. Auch die in einer früheren Monografie beschriebene Signalrotation im Gehirnsystem wird nicht behandelt, da es hier vorrangig um das Wirbeltiergehirn geht, nicht so sehr um das Gehirn des Homo sapiens. Wir müssen uns hier thematisch etwas beschränken.
Über dem analogen Urhirn bildete sich zunächst ein digital arbeitendes Subhirn heraus, über dem sich später ein komplex arbeitendes Subhirn entwickelte. Daher haben moderne Wirbeltiere ein analoges, ein digitales und ein komplexes Subhirn. Der Weg zum digitalen Subhirn führte über die zweite Erweiterungsphase des Spinocerebellums, eine parallele Entwicklung vollzog sich in den Substrukturen der Amygdala. Dies wird nachfolgend beschrieben.
Das komplexe Subhirn arbeitet jedoch nicht mit komplexen Zahlen, wie man vermuten könnte, sondern es arbeitet mit Komplexsignalen. Dieser Begriff wird in der Neurologie bereits seit langer Zeit verwendet und behält in dieser Monografie seine bisherige Bedeutung bei, nur dass diese mathematisch beschreibbar wird.
Gewöhnungsbedürftig ist das digitale Format im Wirbeltiergehirn. Es gibt keine Ziffern. Die Zahlenwerte muss man sich vorstellen wie auf einem Zahlenstrahl mit den natürlichen Zahlen dargestellt, jedoch ist die Zahlenmenge endlich. Jedem Zahlenwert ist ein Outputneuron zugeordnet. Der eigentliche Zahlenwert wird durch die Aktivität einer Neuronengruppe dargestellt, die wir als Extremwertcodierungbezeichnen. Diese ist auf zweierlei Art möglich. Bei der Maximumcodierungwird der Zahlenwert verschlüsselt als Maximum in der aktiven Neuronengruppe, die übrigen Neuronen sind inaktiv. Bei der Minimumcodierungsind dagegen alle Neuronen aktiv mit Ausnahme einer Neuronengruppe, die ein absolutes Erregungsminimum enthält.
Eine solche Codierungsart ist ungewöhnlich, bietet jedoch einen großen Vorteil: Fällt ein Neuron aus, so übernimmt ein Nachbarneuron die Extremwertverschlüsselung. So erreichen neuronale Schaltungen eine unerreichte Ausfallsicherheit.
Eine Übersicht über die strukturelle Dreiteilung des Wirbeltiergehirns in das primäre, das sekundäre und das tertiäre Subhirn liefert die nachfolgende Abbildung.
Nicht eingezeichnet sind die Eingangs- und Ausgangskerne, da sonst die Darstellung unübersichtlich geworden wäre. Am Anfang eines jeden Pfeiles muss man sich einen Ausgangskern vorstellen, dessen Neuronen den Input empfangen und mit ihren Axonen zum Zielsystem leiten. Dort nehmen die Neuronen des Eingangskerns dieser Teilstruktur die Signale entgegen. Bei 13 Teilsystemen gibt es prinzipiell 13 Eingangskerne und 13 Ausgangskerne. Die Signale verschiedener Modalitäten verwenden jedoch meist eigene Teilsysteme. Beispielsweise verarbeiten die verschiedenen Lobi des Gehirns unterschiedliche Modalitäten, ebenso etwa das olfaktorische und limbische System. Dadurch ist die Anzahl der nötigen Neuronenkerne, die als Input- oder Outputkerne fungieren, deutlich größer. Daher sind sie der Übersichtlichkeit zuliebe in der nachfolgenden Übersicht weggelassen worden.
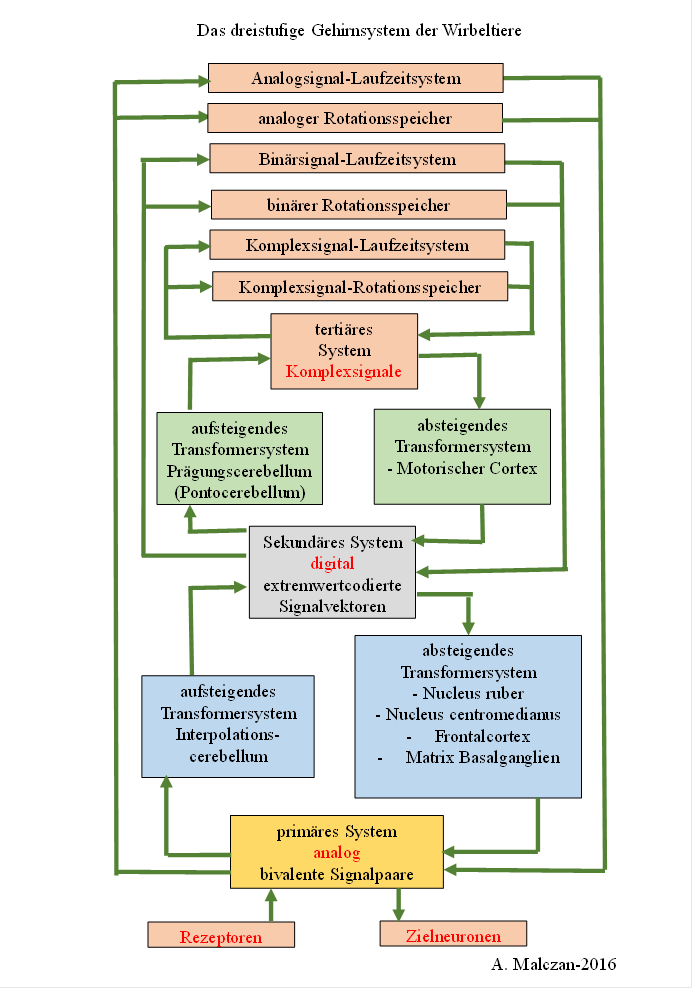
Abbildung 22 - Prinzipschaltbild des dreiteiligen Wirbeltiergehirns nach A. H. Malczan
Alle wesentlichen Komponenten des analogen Urhirns wurden in den bisherigen Kapiteln beschrieben. Nun wenden wir uns dem digital arbeitenden Subhirn zu.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan