Gehirntheorie der Wirbeltiere
für das Human Brain Projekt der Europäischen Union
ISBN 978-3-00-064888-5
3.14 Die Entstehung des urtümlichen Pontocerebellums
Bei denjenigen Bilateria, die ein urtümliches Spinocerebellum hervorbringen konnten, bildete sich parallel dazu ein Pontocerebellum. Dieses bestand anfänglich aus einer extrem geringen Zahl von Neuronen, aber seine Bildung erfolgte gesetzmäßig.
Wir rekapitulieren: Es gab zwei Arten von Outputneuronen im frühen Urhirn dieser segmentierten Bilateria. Einerseits projizierten Neuronen der Klasse 5über den Nucleus ruber abwärts zu den Motoneuronen sowie über den Nucleus olivariszum urtümlichen Spinocerebellum. Diejenigen Neuronen der Klasse 5, die die Vestibularsignale weiterleiteten, projizierten zum Vestibulocerebellum, die anderen ins Spinocerebellum.
Es gab jedoch auch die Neuronenklasse 6der Mittelwertneuronen der ersten, cortikalen Etage. Diese projizierten direkt in das thalamische Mittelwertzentrum(Nucleus subthalamicus) und indirekt über die Projektionsneuronen der Klasse 5 zu den Mittelwertzentren der tieferen Etagen.
Über den Nucleus ruber erreichten die cortikalen Mittelwertsignale auch tiefer gelegene Mittelwertkerne, so dass Letztere ihre segmentbezogenen Mittelwerte durch diese ergänzten und so auch Körpermittelwertebilden konnten. Diese absteigende Projektion erfolgte über den Nucleus ruber.
Hier kam es zur Gleichbehandlung der eintreffenden Signale im Nucleus ruber. Egal, ob es sich um Signale der Vestibularrezeptoren, der Muskelspannungsrezeptoren oder um Mittelwertsignale handelte, alle wurden (im Verlaufe der Zeit) gleichbehandelt: Sie zogen auf den Axonen des Nucleus ruber auch zum Nucleus olivaris. Dessen Axone zogen zum sich bildenden Cerebellum, das anfänglich ein reiner Umschaltkern (Nucleus Purkinje) war und die Signale von Glutamat auf den Transmitter GABA umschaltete. Im Kleinhirnkern erfolgte die Hemmung der dauererregten Outputneuronen, die den erregenden Output an den kontralateralen Nucleus ruber übertrugen. Die Dauererregung entstammte der Formatio reticularis. Dies entsprach einer Signalinversion.
Damit invertierte das urtümliche Cerebellum genau drei Signalarten:
- Vestibularsignale
- Signale der Muskelspannungsrezeptoren (und anderer Rumpfrezeptoren)
- Mittelwertsignale
Gleichartige Signale bildeten im Cerebellum eigene Teilstrukturen. Aus dem Teil, der die Vestibularsignale invertierte, entstand das Vestibulocerebellum. Sein Kleinhirnkern war der Nucleus fastegii.
Der Teil, der anfänglich vor allem die Muskelspannungsrezeptoren invertierte, wurde zum Spinocerebellum. Und da die motorischen Signale von zwei Eingangskernen, dem Nucleus cuneatus und dem Nucleus gracilis kamen, teilte sich der Kleinhirnkern des Spinocerebellums ebenfalls in zwei Teilkerne, den Nucleus emboliformisund den Nucleus globosus.
Der dritte Teil, der die Mittelwertsignale invertierte, wurde zum urtümlichen Pontocerebellum. Sein Kleinhirnkern wurde der Nucleus dentatus.
Es mag vermessen erscheinen, in einer so frühen Phase der Evolution vom Vestibulo-, Spino- und Pontocerebellum zu sprechen. Die genannten Strukturen bestanden aus einer Ansammlung von wenigen Neuronen. Andererseits ist das Pontocerebellum als Ganzes gewiss nicht einfach über Nacht entstanden, sondern muss zwangsläufig eine Evolution durchgemacht haben, die bei einem sehr kleinen Neuronenhaufen begonnen hat. In dieser Monografie wird der weitere Entwicklungsweg aufgezeigt, so wie er sich aus den aktuellen Kenntnissen rekonstruieren lässt.
Theorem des urtümlichen Pontocerebellums
Das urtümliche Pontocerebelluminvertierte die Mittelwertsignale der Klasse 6 der obersten, cortikalen Etage. Sein Output diente der inversen Erregung der kontralateralen Mittelwertsysteme, die es über den Nucleus ruber erreichte.
Über das urtümliche Pontocerebellum tauschten die Mittelwertsysteme ihre Erregungen aus.
Die inverse Erregung der kontralateralen Mittelwertkerne erfolgte topologisch geordnet nach Segmenten.
Wie im frühen Vestibulo- oder Spinocerebellum gab es im urtümlichen Pontocerebellum keine Moosfaserprojektion und keine Körnerzellen. Ebenso gab es keine Sternzellen, Korbzellen oder Golgizellen. Die Cerebellumrinde bestand lediglich aus Purkinjezellen, deren Fläche wir anfänglich als Purkinjekernbezeichnet haben. Deren Dendritenbäume waren anfänglich gewiss klein und rund. Die Herausbildung von hemmenden Interneuronen begann sicherlich damit, dass die laterale Nachbarhemmung zur Kontrastverstärkung zum Standardprogramm in neuronalen Kernen wurde. Solche Interneuronen benötigen jedoch erregenden Signalzustrom, der im urtümlichen Cerebellum noch nicht vorhanden war. Dazu musste sich erst eine erregende Moosfaserprojektion etablieren.
Wir unterstellen, dass die Mittelwertsignale bereits im Nucleus olivaris eine eigene, zusammenhängende Fläche beanspruchten, die jedoch wegen der extrem geringen Signalanzahl nur einen ganz schmalen Streifen bildete, der von der kopfseitigen zur schwanzseitigen Seite verlief und beim bilateralen Lebewesen doppelt vorhanden war. So bekam der wurmförmige Nucleus olivaris, wenn man sich ihn waagerecht angeordnet vorstellt, zwei schmale seitliche Streifen, die die Mittelwertgebietedarstellten, in denen die Axone der Mittelwertneuronen eintrafen.
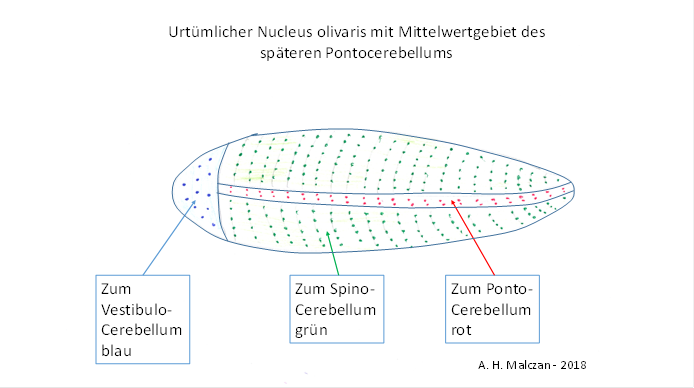
Abbildung14- Urtümlicher Nucleus olivaris
Da diese Neuronen ins Cerebellum projizierten, fand dort die gleiche topologische Veränderung statt. Das Spinocerebellum, waagerecht angeordnet, bildete analog einen länglichen, zeppelinartigen Körper, der zwei seitliche Streifen aufwies. In diesen endeten - für jede Körperhälfte getrennt, die vom Nucleus olivaris eintreffenden Axone mit den cortikalen Mittelwertsignalen. Diese zwei Streifen bildeten das urtümliche Pontocerebellum. An der Spitze dieser Struktur befanden sich die Neuronen, deren Input vom Vestibularsinn stammte und im zugehörigen Vestibulocerebelluminvertiert wurde.
Viele Jahrmillionen später blähte sich diese längliche Struktur des Nucleus olivaris auf, ebenso wie die zugehörige wurmförmige Cerebellumstruktur.
Bezeichnenderweise wird diese Struktur tatsächlich als der Wurm des Cerebellumsbezeichnet, lateinisch als Vermis cerebelli. Deren Mittelwertgebiete bildeten, wenn man sich den Wurm waagerecht angeordnet denkt, anfänglich ebenfalls zwei sehr schmale Streifen, die links und rechts von vorn nach hinten verliefen. Grund war die Tendenz, gleichartige Neuronen zusammenhängend zu platzieren. Vorn befand sich das bilaterale, relativ kleine und abgeflachte Vestibulocerebellum, ebenfalls bilateral links und rechts vorhanden.
Und weil sich die Mittelwertgebiete später deutlich stärker ausdehnten als die zentralen, zum Cerebellumwurm gehörenden Anteile, entstand seitlich links und rechts von diesen das bilateral vorhandene Pontocerebellum. Da es sich dabei genau um die Mittelwertgebiete handelt, sind wir berechtigt, diese frühe Vorläuferstruktur im urtümlichen Spinocerebellum bereits hier als Pontocerebellum zu bezeichnen.
Parallel zur Bildung des urtümlichen Pontocerebellums erfolgte im frühen Urhirn der Bilateria, deren Linien zu den Wirbeltieren führten, eine Gleichbehandlung der Signalklassen5 und 6 im frühen Basalgangliensystem. Ebenso wie die Signale der Klasse 5zogen die Signale der Klasse 6- über den Zwischenkern des Nucleus subthalamicus- absteigend zum Nucleus ruber. Von dort zogen die Signale der Klasse 5 zu den Motoneuronen, aber die der Klasse 6 zu den verschiedenen Mittelwertkernen.
Theorem der Projektionen des Nucleus subthalamicus
Der Output des Nucleus subthalamicus erreicht absteigend einerseits den Nucleus ruber, andererseits auch die Substantia nigra pars compactasowie über das Cerebellum auch die Formatio reticularis.
Zwischen den Mittelwertsystemen der verschiedenen Etagen gab es einen Signalaustausch, wobei der Nucleus subthalamicus das ranghöchste System war. Er projizierte auch erregend in die dopaminerge Substantia nigra pars compacta.
Da jedes Mittelwertsystem erregend in die ursprünglichen Herkunftsgebiete des Inputs zurückprojiziert, gibt es auch eine aufsteigende Projektion von der Substantia nigra pars compacta kopfwärts. Die vom Nucleus subthalamicus stammenden Mittelwertsignale der cortikalen Cluster wurden in der Substantia nigra pars compacta auf Dopaminumgeschaltet und zurückgeschickt, sie endeten im Striatuman GABAergen Neuronen, die als Striosomenbezeichnet werden. Die Striosomenneuronen werden von diesem Input erregt, da sie den Dopaminrezeptor D2verwenden.
Dies führte - bedingt durch die relativ große Entfernung zwischen beiden Strukturen - zu einer Zeitverzögerung der Striosomensignale. Der Output der Striosomenneuronen der Klasse 6erreicht über den Tractus tegmentalis centralis den Nucleus ruber. Dort gibt es bereits signalverwandte Neuronen, die vom Nucleus subthalamicus ebenfalls das Mittelwertsignal eines zugeordneten Cortexclusters empfangen. An diesen Neuronen docken die striatalen Axone der Striosomen der Klasse 6 an und hemmen diese mit zeitlicher Verzögerung.
Damit entstand im Nucleus rubereine zeitsensitive Differenzabbildungder kortikalen Signalmittelwerte. Nun konnte das Striosomensystem sowohl zeitliche Veränderungen in der Signalklasse 5 erkennen als auch solche in der Signalklasse 6.
Theorem der zeitsensitiven Differenzabbildung der Signalmittelwerte im Nucleus ruber und des Kletterfaserinputs des Pontocerebellums
Neben der Signalklasse 5wurde auch die Signalklasse der Mittelwertsignale in das Striosomensystemeingebunden und ließ im Nucleus ruber eine zeitsensitive Differenzabbildung der Signalmittelwerte entstehen.
Die Gleichbehandlung der Signaleim Nucleus ruber führte dazu, dass diese Outputsignale der Mittelwert-Differenzabbildung über den Nucleus olivaris das Cerebellum erreichten. Da sie der Signalklasse 6 zugeordnet waren, erreichten diese Signale auf den Axonen der Kletterfaserndie Purkinjezellendes sich bildenden Pontocerebellums.
Damit konnte das urtümliche Pontocerebellum zeitliche Veränderungen in den Signalmittelwertender verschiedenen Segmente des Körpers erkennen und aus ihnen eine inverse Erregungder kontralateralen Segmentanteile ableiten. So konnten in den einzelnen Segmenten Urformen von Taktgeneratorenentwickelt werden, die eine asynchrone, gegensätzliche Ansteuerung der Motorik gestatteten und eine schlängelnde Fortbewegung ermöglichten, wie sie noch heute bei Fischen zu beobachten ist.
Die Striosomen können wir nun ebenfalls in zwei Unterarten einteilen. Ein Teil verarbeitet die Signale der Signalklasse 5, der andere die der Klasse 6. Dies berechtigt uns dazu, von Striosomen der Klasse 5 und 6 zu sprechen.
Theorem der Klassifizierung der Striosomen
Die Striosomen der Klasse 5verarbeiten Signale der Klasse 5 und dienen der Bildung einer zeitsensitiven Differenzabbildung der Muskelspannungsrezeptorenim Nucleus ruber, der diese sowohl den Motoneuronen als auch über den Nucleus olivaris dem Spinocerebellum zustellt. Im Vestibulocerebellum erlaubt sie die Erkennung von Veränderungen der Körperlage im Gravitationsfeld der Erde, die im Vestibulocerebellum ausgewertet werden. Im Spinocerebellum gestattet sie die Erkennung von Signalveränderungen der verschiedenen Modalitäten und somit die Bewegungserkennung mittels verschiedener Rumpf- und Kopfsinne.
Die Striosomen der Klasse 6verarbeiten die Mittelwertsignale der Klasse 6 und dienen im Nucleus ruber der Bildung einer zeitsensitiven Differenzabbildung der Mittelwertsignaledes Systems. Sie erlaubt die Erkennung von zeitlichen Veränderungen in der Signalstärke der Mittelwertsysteme. Der Nucleus ruber leitet diesen Output weiter sowohl an andere Mittelwertsysteme als auch über den Nucleus olivaris an das Pontocerebellum.
Die Axone von den Striosomen zum Nucleus ruber bilden bei höheren Säugetieren einen markanten Nervenstrang, der als Tractus tegmentalis centralisbezeichnet wird. Anfänglich, als der Körper der Tiere nur wenige Muskeln besaß, war er äußerst unscheinbar, die Anzahl der beteiligten Neuronen war zunächst sehr gering. Dennoch soll dieser Neuronenstrang bereits hier mit diesem Namen bezeichnet werden.
Mit der Einbeziehung der Signalklasse 6 in die Striosomen des Basalgangliensystemsgalt es allerdings, ein damit verbundenes Problem zu lösen. Die Signalklasse 6 entsprach den cortikalen Mittelwerten. Nun gibt es genügend cortikale Gebiete, die ununterbrochen mit Input versorgt werden. Denken wir nur an die Sehrinde. Diese empfängt am Tage ständig Signale der Sehrezeptoren der Netzhaut. Damit sind die daraus gebildeten Mittelwertsignale dauerhaft stark und liefern ein starkes Mittelwertsignal an die Substantia nigra pars compacta. Diese projiziert erregend zum Striatum.
Eine ständige und starke Dauererregung ist für Neuronen jedoch tödlich. Daher war es zweckmäßig, diese zu dämpfen. Die Lösung war simpel und naheliegend: Die vom Striatum abwärts ziehenden Axone (der Mittelwertsignale) kamen im Nucleus ruber an und bewirkten dort die Bildung der zeitsensitiven Differenzabbildung. Doch sie setzten ihren Weg fort und erreichten ebenso die Substantia nigra pars compacta. Dort wirkten sie wegen ihres Transmitters GABA hemmend und verhinderten eine ununterbrochene Dauererregung. Da es sich um starke Mittelwertsignale der Klasse 6 handelte, war deren Hemmungswirkung stark und stoppte die Dauererregung in der Substantia nigra pars compacta für etliche Millisekunden, bis die GABAergen Signale aus dem Striatum aufgebraucht waren. Dann fiel diese Hemmung weg und die Substantia nigra wurde wieder von den cortikalen Mittelwertsignalen erregt, die ja ständig diesen Kern erreichten. Daher konnte die Substantia nigra wiederum das Striatum, genauer dessen Striosomen erregen. Diese projizierten aber wieder hemmend in diesen Kern zurück, so dass - wegen der langen Wege - die hemmenden Aktionspotentiale nach einer Weile diesen Kern wieder hemmten.
So wurde der Output der Substantia nigra von regelmäßigen Hemmungspausen unterbrochen. Die Signalklasse 6 wurde dadurch in ein tonisches (höherfrequentes) Signal transformiert, welches regelmäßige Pausen aufwies.
Genau dies ist das sogenannte Kletterfasersignal, welches an den Purkinjezellen des Pontocerebellums nachgewiesen wurde. Es liegt genau dann an diesen Purkinjezellen an, wenn das zugeordnete cortikale Mittelwertneuron der Klasse 6 aktiv ist, weil im zugehörigen Einzugsbereich dieses Mittelwertneurons die Cortexneuronen erregt werden.
Theorem der hemmenden Rückkopplung von den Striosomen des Striatums zur Substantia nigra pars compacta
Die Projektion der cortikalen Mittelwertneuronen der Klasse 6 in die Substantia nigra pars compacta erforderte eine Erregungsbegrenzung. Daher etablierte sich eine hemmende Projektionvon den Striosomenneuronen des Striatums in die Substantia nigra pars compacta. Dadurch wurde erreicht, dass diese Neuronen nicht dauerhaft und pausenlos erregt waren, sondern ihre tetanische Schwingung von kurzen Hemmungpausenunterbrochen wurde, in denen sich die Neuronen reaktivieren konnten. Die Ursache für die Hemmungspausen war die Laufzeit der Signale zwischen Cortex und Substantia nigra pars compacta. Diese Hemmungspausen wurden auch den Striosomenneuronen der Klasse 6aufgeprägt. Deren Outputsignale hemmten im Nucleus ruber die zugehörigen tetanischen Signale der Klasse 6, so dass die Outputsignale dieser Neuronen des Nucleus ruber ebenfalls tetanische, aber von Pausen unterbrochene Schwingungenwaren. Dieser Output erreicht über den Nucleus olivaris das Pontocerebellum als Kletterfasersignale.
Die Zeitverzögerungerklärt die Besonderheit der Kletterfasersignale im Pontocerebellum, die darin besteht, dass die höherfrequente (tetanische) Schwingung von kurzen Pausen unterbrochen wird, die für den Lernprozess eine wichtige Bedeutung haben.

Abbildung15- Im Striosomensystem entstehendes Kletterfasersignal
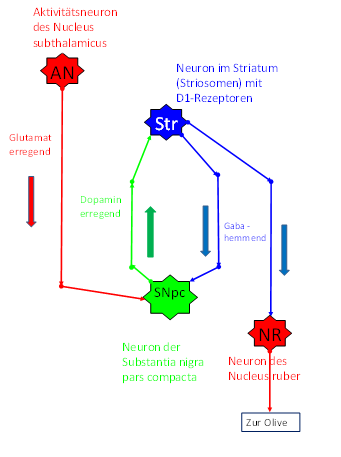
Abbildung16- Dopaminerge und GABAerge Projektion im Basalgangliensystem
Die Zeitverzögerung der dopaminergen Signaleauf dem Weg von der Substantia nigra pars compacta zum Striatum und der GABAergen Signale vom Striatum zurück zur Substantia nigra pars compacta führt zu periodischen Unterbrechungder Mittelwertsignale. Damit entsteht durch das Striosomensystem der Klasse 6 eine charakteristische Schwingungsform, die wir (in dieser Monografie) als Kletterfasersignal bezeichnen. Jeder einzelne Cortexcluster liefert auf diese Weise ein eigenes Kletterfasersignalzu seinem Cerebellumcluster im Pontocerebellum. Da der Begriff des Cortexclusters bisher nicht erklärt wurde, sei der Kürze halber vermerkt, dass ein Cortexcluster das Einzugsgebiet eines cortikalen Mittelwertneurons darstellen möge. Darauf wird später genauer eingegangen.
Bereits auf dieser frühen Stufe der Evolution wurden so Voraussetzungen für die spätere Lernfähigkeit des Pontocerebellumsgeschaffen. Die für eine Langzeitpotenzierung (LTP) oder Langzeitdepression (LTD) notwendige hochfrequente (tetanische) neuronale Schwingung wird im Striosomensystem der frühen Basalganglienerzeugt. Dies geschah bereits zu einer Zeit, als das Pontocerebellum sich gerade herauszubilden begann.
Theorem der striosomalen Kletterfasersignale des Pontocerebellums
Die striosomalen Kletterfasersignaledes urtümlichen Pontocerebellums waren von kurzen Pausen unterbrochene tetanische Schwingungen, die für die später auftretende Langzeitdepression (LTD) und Langzeitpotenzierung (LTP)im Pontocerebellum unabdingbar waren. Immer dann, wenn in einem beliebigen Cortexcluster eine neuronale Erregung auftrat, wurde diese erregend vom zuständigen Mittelwertneuron sowohl zum Nucleus subthalamicus als auch zur Substantia nigra pars compacta gesendet und stieß eine getaktete Dauerschwingung an, die wir als Kletterfasersignal bezeichnen. Dieses wurde durch negative Rückkopplung periodisch unterbrochen, damit keine Überlastung der beteiligten Neuronen auftrat. Über den Tractus tegmentalis centralis erreichte das Striosomensignal der Klasse 6den Nucleus ruber und hemmte das vom Nucleus subthalamicus eintreffende Mittelwertsignal der Klasse 6 in einer Punkt-zu-Punkt-Abbildung, so dass diesem Austastpausen aufgezwungen wurden. Dieses getaktete Kletterfasersignal eines cortikalen Clusters erreichte über den Nucleus olivaris und letztlich über die Kletterfasern die Purkinjezellen des sich bildenden Pontocerebellums, ebenso auch die Outputneuronen des Nucleus dentatus. Durch die Existenz der tetanischen Schwingungen konnte sich im Pontocerebellum ein Lernalgorithmusherausbilden, der auf dem Phänomen der Langzeitdepression(LTD) und später auch auf der Langzeitpotenzierung(LTP) beruhte.
Der Lernalgorithmus wird in nachfolgenden Kapiteln beschriebenen. Er konnte sich erst herausbilden, als die zweite Erweiterungsphase des Pontocerebellums begann, in der sich cerebellare Interneuronen und ein Signalzustrom über ein Moosfasersystem herausbilden konnte.
Wichtig für das Verständnis der Lernvorgänge im sich entwickelnden Pontocerebellum ist die Synchronität der Schwingungsvorgängeim Striosomensystem der Basalganglien, die sich sowohl in der Substantia nigra pars compacta als auch in den Striosomen und im Nucleus subthalamicus bemerkbar macht. Zilles und Rehkämper beschreiben in [5] auf Seite 287 die schrittmacherartige Aktivitätden großen, dopaminergen Neuronen in der Substantia nigra pars compacta.
Theorem der Synchronität der Schwingungsvorgänge im Striosomensystem der Basalganglien
In der Substantia nigra pars compactawerden die Schwingungspausen zwischen den vielen beteiligten Neuronen synchronisiert.
Theorem der Striosomen als Mittelwertrepräsentanten der Cortexcluster
Da jedes Neuron der Klasse 6 den Signalmittelwert eines cortikalen Clusters repräsentiert, der über die Substantia nigra pars compacta auf Dopamin umgeschaltet wird und ein Striosomenneuon der Klasse 6 im Striatum erreicht, repräsentiert auch jedes dieser Striosomenneuron den Signalmittelwert eines zugeordneten cortikalen Clusters. Die Mittelwertschwingungen werden durch synchronisierte Pausenunterbrochen.
Es sei daran erinnert, dass es bereits ein Striosomensystem im Urstriatum gab, in dem die Signale der Signalklasse 5ebenso verarbeitet wurden wie nun die Signale der Signalklasse 6. Wir können diese Striosomenneuronen unterscheiden, indem wir ihnen die Signalklasse zuordnen, die sie verarbeiten. Dann gibt es Striosomenneuronen der Klasse 5und der Klasse 6. Letztere zählen zum Mittelwertsystem. Beide Arten werden vom dopaminergen Input erregt.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan