Gehirntheorie der Wirbeltiere
für das Human Brain Projekt der Europäischen Union
ISBN 978-3-00-064888-5
3.10 Theorie der neuronalen Signalinversion
Wir wenden uns zunächst den Mittelwertkernenzu, die in etlichen Segmenten der segmentierten Bilateria ihr Werk verrichten. Bisher gingen wir davon aus, dass die Signalmittelwerte der verschiedenen Modalitäten zur Steuerung der Lebensfunktionen verwendet werden. Die mittlere Helligkeit steuert beispielsweise den circadianen Rhythmusdes Lebewesens (innere Uhr). Der Mittelwert der motorischen Aktivität sagt etwas aus über den damit verbundenen Energieverbrauch und die nötige Energiebereitstellung. Der mittlere Wert der olfaktorischen Erregung erlaubt die Einschätzung darüber, ob fressbare Beute in der Nähe ist. Viele andere Prozesse sind ebenfalls in Regelkreise eingebunden, die Signalmittelwerte benötigen.
Weiterhin wenden wir uns den hemmenden Neuronenzu. Zunächst werden sie als hemmende Interneuronen in den sensorischen und motorischen Zentren des Strickleitersystems für die laterale Hemmung verwendet, mit welcher der Kontrast zwischen starken und schwächeren Signalen verstärkt wird.
In Neuralrohrsystem erfüllen sie in den verschiedenen Segmenten und Ebenen die gleiche Funktion. Hierbei ist die Hemmung oft relativ. Ein stark erregtes hemmendes Neuron hemmt stark, ein schwach erregtes nur schwach. Eine totale Hemmung unabhängig von der aktuellen Feuerrate wäre für die laterale Hemmung unbrauchbar. Die Hemmungswirkung ist von der aktuellen Feuerrate abhängig und nimmt mit ihr zu.
Wir verknüpfen nun die Mittelwertbildung mit der relativen Hemmung, um ein Signal S zu invertieren. Aus Gründen, die erst im Kapitel 4 dieser Monografie genauer erklärt werden, bilden wir aus einem Signal mit der Feuerrate f und einem Mittelwertsignal mit der Feuerrate fm ein invertiertes Signalf*, wobei die Verknüpfung nicht additiv, sondern multiplikativ ist und der Formel
![]() (1.8.1)
(1.8.1)
genügt. Daraus folgt unmittelbar
![]() (1.8.2)
(1.8.2)
Das Produkt einer Feuerrate und ihrer invertierten Feuerrate ist gleich dem Quadrat der Feuerrate des Mittelwertes.
Wenn man davon ausgeht, dass Rezeptoren und Neuronen eine exponentielle Kennliniehaben, führt das Logarithmieren auf die ursprünglichen Größen, von denen die Erregung abgeleitet wird. Insofern ist es nicht verwunderlich, dass zwischen einer Feuerrate f und ihrer Invertierten f* die Beziehung
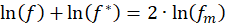
besteht. Insofern kann man die Signalinversionals logarithmische Mittelwertspiegelungbezeichnen.
Theorem der Signalinversion
Ein Signal mit der Feuerrate f wird invertiert, indem das Quadrat der mittleren Feuerrate fm durch die Feuerrate f dividiert wird.
Realisiert wird die Signalinversion dadurch, dass das Signal mit der Feuerrate f zunächst auf einen hemmenden Transmitter umgeschaltet wird - falls es nicht selbst hemmend ist - und einem Outputneuron zugeführt wird, welches gleichzeitig von einem Mittelwertsignal mit der Feuerrate fm erregt wird. Durch die nichtlineare, exponentielle Kennlinie dieses Outputneurons entsteht durch die Überlagerung von Erregung und Hemmung das invertierte Signal f*.
Eine Schaltung, die ein Signal invertiert, bezeichnen wir als (neuronale) Inversionsschaltung.
Besitzt das Signal S an einer Stelle ein Minimum, so besitzt das invertierte Signal S* dort ein Maximum. Die Monotonie wird durch die Inversionstransformationumgekehrt.
Theorem der Signalinversion von minimumcodierten Signalen
Ein minimumcodiertes SignalS wird durch eine Inversionsschaltung in ein maximumcodiertes Signalüberführt.
Für die genauere Identifizierung von Neuronen in Inversionsschaltungen führen wir folgende Bezeichnungen ein. Das Inputneuronstellt das erregende Signal bereit, welches invertiert werden soll. Das nachfolgende, GABAerge Neuron bezeichnen wir als Umschaltneuron, weil hier die Transmitterumschaltung beispielsweise von Glutamat auf GABA erfolgt. Das Outputneuron bezeichnen wir als Inversionsneuron, weil es unter Verwendung des zugeführten Signalmittelwertes und des hemmenden Umschaltneurons die Signalinversion vornimmt. Ein Mittelwertneuronführt dem Inversionsneuron den nötigen Signalmittelwert zu, damit eine Inversion überhaupt stattfinden kann.
Die Verwendung anderer hemmender Transmitter ist durchaus möglich.
Eine Signalinversion überführt ein starkes Signal in ein schwaches und umgekehrt, eine Monotonie im Signalverlauf kehrt sich um.
Häufig werden die Signale vieler Neuronen invertiert, die alle auf einer Linie oder auf einer Fläche verteilt sind. In diesen Fällen muss nicht jedes der zugehörigen Inversionsneuronen ein eigenes, separates Mittelwertneuron in Anspruch nehmen, sondern ein Mittelwertneuron kann seinen Signalmittelwert gleichzeitig auf viele Inversionsneuronen verteilen, indem sich sein Axon entsprechend häufig verzweigt. Der Signalmittelwert kann auch aus einem weiter entfernten Mittelwertkern abgezapft werden.
Bei einer realen Signalinversion kommen sicherlich nichtlineare Abweichungenvon der Idealformel vor. Dies ist unbedeutend, solange die Monotonieumkehrerfolgt, also Minima zu Maxima werden und Maxima zu Minima. Nötig ist also vorrangig die Extremwertumkehr. Diese könnte durch vielerlei neuronale Kennlinien von Inversionsneuronen realisiert werden.
Mit der Signalinversion erhielten die hemmenden Neuronen eine neue Aufgabe: die relative Hemmung von Mittelwertsignalenzur Transformation eines Signals in sein invertiertes Signal. Diese Transformation wird zum Arbeitsprinzip des sich bildenden Vestibulocerebellums.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan